
記住我
Serotonin (5-hydroxytryptophan, 5-HT) is a monoamine neurotransmitter that regulates many physiological functions [1], and is phylogenetically conserved in several species, including humans, mice, and fruit flies. Serotonin is synthesized from the diet-derived amino acid tryptophan, which travels throughout the blood to act as a clotting factor with wound healing. Remarkably, most serotonin is located in the gut in enterochromaffin cells, although correlations of gut serotonin and depression are not well established. Brain serotonin is created from tryptophan, which crosses the blood-brain barrier and enters the dorsal raphe nuclei (DRN) of the brain stem [49]. Primarily, serotonin cell bodies radiate from the DRN to the substantia nigra, hypothalamus, nucleus accumbens, and medial prefrontal cortex [49, 50], and these diverse signaling pathways allow serotonin to regulate different behaviors like sleep, mood, memory, and appetite [50, 51]. The serotonin transporter (SERT) provides a negative feedback loop to reuptake extracellular serotonin back into the neuron to clear released serotonin, which is shown in Fig. 5A [1, 50]. After reuptake, excess serotonin is either re-packaged into vesicles for release again or destroyed by lysosomes [52]. Additionally, there are seven classes of serotonin receptors in mammals that regulate serotonin neuronal activation and inhibition [1, 50]. All are ligand-gated ion channel GPCRs, except 5-HT-3 receptors. The GPCR serotonin receptors are also illustrated in Fig. 5A [1, 50]. These receptors act as either autoreceptors or heteroreceptors. Autoreceptors are located on presynaptic neurons and attenuate serotonin release with a negative feedback loop, while heteroreceptors are on post-synaptic neurons and regulate serotonin through postsynaptic feedback [1, 50]. In addition to mammals, Drosophila melanogaster also possess SERT (dSERT) and serotonin receptors that show homology with structure and function [53,54,55]. Although a controversial new article suggests that serotonin is not involved in depression, decades or work have shown changes with serotonin with both genetic manipulations and antidepressants [56]. Specifically, these bioanalytical techniques that measure serotonin help identify how SERT and serotonin receptors control serotonin signaling in the brain, which gives insight into how their dysfunction causes depression, anxiety, and aggression.
Fig. 5
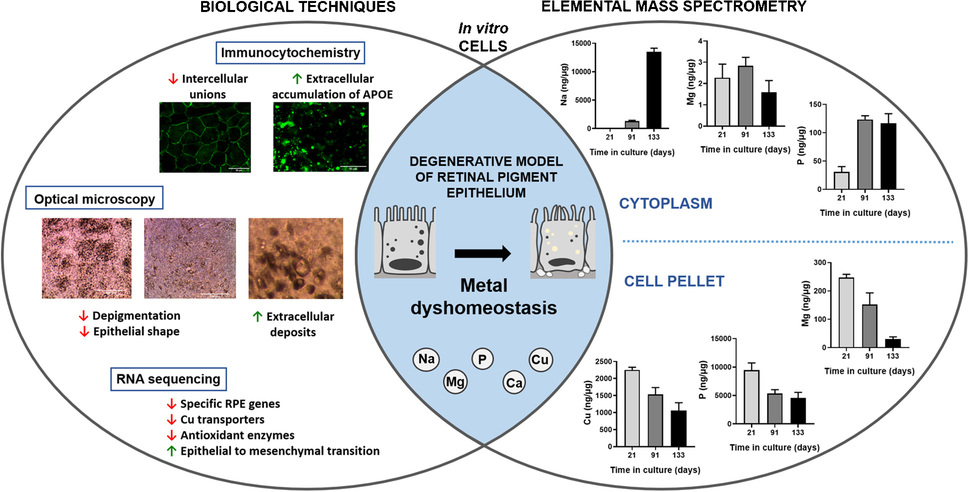
Electrochemical techniques show serotonin uptake changes with different SSRI antidepressants. A (i) Schematic of serotonergic system with serotonin transporters (SERTs) and serotonin receptors. Long (l) and short (s) allele variant for 5-HTTLPR and its theoretical changes to serotonin are also listed. (ii) Chronoamperometry was used to show lymphocytes (immune cells) from rhesus macaques can uptake serotonin and paroxetine inhibits uptake. (iii) (s)-Allele 5-HTTLPR shows faster reuptake of serotonin in lymphocytes compared the (l) variant (permission from Singh et al. 2010, ACS Chemical Neuroscience). B Schematic of the design and application of an aptamer-FET biosensor for in vivo serotonin measurement (permission from Zhao et al. 2021, Science advances). C (i–iii) FSCV representative fast, slow, and hybrid serotonin kinetic responses from stimulated serotonin release in the substantia nigra (permission from Wood et al. 2014, Journal of Neurochemistry). Data were used to create the free kinetic modeling program, The Analysis Kid, using FSCV. D FSCV data shows serotonin concentration and reuptake changes with a low, 1 µM dose of escitalopram, citalopram, paroxetine, and fluoxetine in fruit flies. Each SSRI changed serotonin differently and concentration and reuptake are coupled for some, but independent for others (permission from Dunham and Venton 2022, Journal of Neurochemistry)
With chronoamperometry, the Andrews lab explored several fundamental changes to the serotonin system with genetic mutations and antidepressants [57, 58]. For example, they explored the effect of a 50% SERT knockdown (SERT +/-) in mice versus a complete SERT knockout (SERT-/-) on serotonin clearance rates in brain liposomes or synaptosomes [57]. Serotonin clearance rates increased with increased oxygen. Also, decreased SERT expression increased uptake of serotonin, which was confirmed with paroxetine (1 µM) treatments in control SERT +/+ mice [57]. Likewise, serotonin uptake rate changed with a short-allele SERT variant in rhesus macaques. In mammals, a short (s) 5-HTTLPR allele variant decreased mRNA and protein expression of SERT, while the long (l) allele did the opposite, and changed basal serotonin concentrations (Fig. 5A) [58]. They used a novel boron-doped diamond microelectrode, which decreased electrode fouling to serotonin with chronoamperometry. Additionally, lymphocytes actively cleared serotonin and serotonin uptake was inhibited by SSRIs, like paroxetine (Fig. 5Aii). The (s) 5-HTTLPR allele showed faster serotonin clearance compared to the (l) allele variant, and the homozygous (s) allele had faster reuptake of serotonin than a heterozygous allele (Fig. 5A). Together, these data suggest that the (s) 5-HTTLPR allele variant reuptake more quickly, which may alter serotonin release mechanisms differently to maintain higher basal serotonin concentrations. Recently, the Andrews lab has also created an implantable aptamer-FET neuroprobe biosensor that measures serotonin changes in mice in vivo, which is illustrated in Fig. 5B [59]. Since this technique is very new, it has not been characterized extensively, but it will allow for the real-time measurement of serotonin to understand depression behavior and treatments in the future. Thus, analytical techniques for measuring serotonin have shown different serotonin levels with different serotonin genotypes, which gives insight into why some patients may respond differently to different antidepressants.
Also with chronoamperometry, the Daws group studied serotonin clearance and antidepressant mechanisms [31, 33, 60]. Like many fundamental studies of serotonin uptake, these studies were performed in anesthetized animals, which are easier to study than freely moving animals and have similar clearance mechanisms, even under anesthesia. They first characterized fundamental uptake changes with SERT and the norepinephrine transporter (NET) in mice using fluvoxamine and citalopram, and found that both SERT and NET contribute to the active clearance of serotonin in the CA3 region of the hippocampus, which is a very important region studied in depression [9, 60]. They later expanded on this work to explore reuptake serotonin kinetics that were first described by Shashkan and Snyder in the 1970s, specifically “uptake-1” and “uptake-2” of serotonin [61, 62]. Uptake-1 is high affinity for serotonin with slow transport kinetics, while uptake-2 is low affinity with faster transport kinetics. Ultimately, the Daws group used pharmacology to show that low affinity transporters, like organic cation transporters (OCT) and plasma membrane monoamine transporters (PMAT) play important roles in serotonin reuptake [33, 61, 63]. This is an important finding, as it shows that clearance is not just SERT dependent, and that other transporters may be targets for antidepressants as well.
The Daws lab also investigated genetic and modulatory changes to SERT with serotonin receptors and antidepressants [30, 31, 64, 65]. Initially, they explored serotonin changes with male 5-HTT (SERT) KO mice (+/+, +/-, and -/-) [31], and found that serotonin clearance decreased in the heterozygous (+/-) group, and even more in the homozygous (-/-) mice. The SSRI fluvoxamine both inhibits reuptake of serotonin and increases concentrations through SERT, since the (-/-) mice did not change serotonin clearance or concentrations. Likewise, with escitalopram and SERT KO mice, radio-ligand binding assays of [H3]-citalopram show SERT is required for antidepressants to function [65]. They also examined SERT knockout (-/-) and 5-HT1B autoreceptor genotypes [30], and chronoamperometry showed SERT activity did not change with different 5-HT1B mutations in the CA3 region of the hippocampus. However, they found that the 5-HT1B receptor antagonist cyanopindolol did not inhibit serotonin in the 5-HT1B (-/-) or SERT (-/-) mice, which was different from the wild type and suggests that both work together to regulate serotonin inhibition. Ultimately, analytical methods here add functional information about how SERT and 5-HT1B work together in various genotypes with SSRIs.
With FSCV, the Hashemi lab simultaneously measured serotonin and dopamine changes in vivo in rats to understand their dynamic changes in clearance with reuptake inhibitors, as well as vesicular monoamine transporter (VMAT) inhibitors [66]. Dopamine release was more sensitive to changes in vesicular packaging, while serotonin was tightly controlled by SERT through reuptake. Michaelis-Menten kinetics changes after SSRI antidepressants, citalopram and escitalopram, were also similar to previous work by Shaskan and Snyder and the Daws group [67, 68]. The Hashemi lab also created classical models for uptake-1 (fast) and uptake-2 (slow) with antidepressants. However, they saw serotonin traces with escitalopram show “hybrid” clearance of serotonin that starts fast and then slows at low, antidepressant doses [62]. These uptake responses are illustrated in Fig. 5C with FSCV color plots, concentration versus time plots, and release and fraction retained rates. They also used a form of FSCV, called fast-scan cyclic adsorption voltammetry, in order to measure basal levels of serotonin in mice, and saw that females typically have lower concentrations of serotonin compared to males [68]. Further, their models are now available in a web-based tool, known as the Analysis Kid [69], with their previously collected mathematical models for serotonin and dopamine. This software allows users to directly upload current vs. time trace files or color plots to determine Michaelis-Menten kinetic variables for their data. It is important to note that dopamine has two types of uptake (fast and slow), while serotonin has three (fast, slow, and hybrid). Also, even though their models were created with mammals, Drosophila and humans show nearly identical SERT translocation speeds, which would produce similar uptake changes [70].
In Drosophila, the Venton group pioneered using optogenetics, which are light-activated channels specifically expressed in certain cells, to selectively release serotonin that is measured with FSCV [71, 72]. They found the releasable pool of serotonin in Drosophila takes 2–5 min to replenish itself [71, 73], and both serotonin synthesis and repackaging from reuptake by Drosophila SERT (dSERT) were necessary to replenish the releasable pool [73]. Real-time serotonin concentration and reuptake changes were also characterized with different doses of fluoxetine, escitalopram, citalopram, and paroxetine that were bath-applied to Drosophila ventral nerve cord (VNC) tissue [74]. They found SSRIs differentially modulated serotonin reuptake and release based on different dSERT affinities. Specifically, paroxetine showed the highest affinity to dSERT, while fluoxetine the lowest. With escitalopram and citalopram, the S-enantiomer showed higher affinity to dSERT, which caused serotonin concentrations to increase and reuptake to slow at lower doses compared to citalopram. With these SSRIs, reuptake was independent of serotonin release. For instance, at low doses, fluoxetine only slowed serotonin reuptake, but did not increase concentration, while paroxetine showed high concentration increases with fast serotonin clearance. These real-time serotonin changes with low SSRI doses are shown in Fig. 5D. Ultimately, this work shows that Drosophila is a good model for rapid pharmacology screenings to decipher how neurotransmitters change [74, 75].
Together, these analytical techniques provide valuable insights into how serotonin changes with genetic mutations to the serotonin system and with SSRI antidepressants. Fundamentally understanding how serotonin changes with different SSRIs [74], or SERT and serotonin receptor mutations [31, 58, 65], will aid in deciphering an individual’s genetic pre-disposition to react to a specific antidepressant with predictive genomics [76]. Further, the tools created by the Hashemi group allow FSCV users to track kinetic changes with any drug that impacts serotonin or dopamine [69], which opens the field to new uptake models in the future, including the transporters (OCT, PMAT) that have been implicated with antidepressant mechanisms and behavior responses [61, 63]. Likewise, the advent of new, flexible and implantable biosensors will allow for the chronic measurement of serotonin and its relationship to antidepressants and behaviors that will expand our fundamental knowledge of long-term changes in the near future [59].
Histamine, inflammation, and immune cell signalingHistamine (HA) is another monoamine neurotransmitter, similar to serotonin, that plays important roles in sleep-wake cycles, as well as memory formation and learning [11, 77]. However, it also acts as a central signaling molecule that leads to a cascade of cellular immunological responses that cause smooth muscles to contract and blood vessels to become permeable. Histamine increases inflammation and swelling with the activation of radical oxygen species (ROS), which are known to act as a signaling molecules and damage cells. Four sub-classes of histamine receptors (HR1-4) have been identified in mammals, all GPCRs, but they differ in their tissue expression and actions [77]. For instance, HR1 and HR2 receptors are mainly expressed in airways with vascular smooth muscle cells to activate or inhibit the release of histamine from immune cells. HR4 is found in bone marrow and white blood cells. HR3 receptors act as pre-synaptic receptors in the central nervous system and control the release of histamine and other neurotransmitters, such as serotonin, dopamine, norepinephrine, acetylcholine, and GABA [77]. HR3 receptors also connect neurons with immune cells, like mast cells. In addition to mammals, Drosophila melanogaster also produces histamine and possesses homology to these histamine receptors [78]. Histamine activates inflammation and immunological signaling and may regulate other neurotransmitters, like serotonin, so recent studies have concentrated on measuring both serotonin and histamine to determine their roles in depression.
With FSCV, the Hashemi lab developed electrochemical methods to investigate inflammation with depression. They co-detected histamine and serotonin with FSCV using the Jackson waveform in vivo in mammals with Nafion-coated electrodes (Fig. 6A) [40]. They found serotonin and histamine concentrations change with long-term stress and treatment with escitalopram [79], and that stress to mice increased histamine concentrations, while serotonin decreased. Escitalopram alone did not bring serotonin back to baseline levels in stressed mice, but concentrations did increase with combined escitalopram and α-fluoromethylhistidine (FMH), a histidine decarboxylase inhibitor. These results suggest that histamine clearance influences serotonin release and clearance during stress. The Hashemi group also collaborated with the Orian lab to test a novel selenofluoxetine derivative [80]. The selenofluoxetine derivative binds to SERT similarly as fluoxetine to inhibit serotonin reuptake with FSCV and models show that the selenium group adsorbs oxygen radicals that cause inflammation, which are mediated by histamine. Altogether, these analytical tools help us understand real-time serotonin and histamine signaling during stress and with antidepressants and contribute to our basic knowledge of the mechanisms of new antidepressant pharmaceuticals that decrease inflammatory responses.
Fig. 6
Real-time electrochemical techniques show histamine (HA) changes 5-HT. A With stress, increased histamine (i) causes 5-HT to decrease (ii) (permission from Hersey et al. 2022, Journal of Neuroinflammation). B Increased LPS signals for IL-6 and changes serotonin compared to the control (i). Increased LPS decreases serotonin concentration and uptake (ii) over time (iii), but returns to baseline (permission from Zhu et al. 2010, Neuropsychopharmacology). C The Daws group used chronoamperometry to measure 5-HT changes when immepip (H3/H4 receptor agonist) and thioperamide (H3/H4 receptor inverse agonist) were exogenously applied for 30 min to the mice hippocampus. H3/H4 receptor agonism slows serotonin clearance, while H3/H4 receptor inverse agonism both decreases 5-HT concentration and quickens uptake (permission from Annamalai et al. 2020, ACS Chemical Neuroscience). D Example of an IL-6 cytokine aptamer-based biosensor that could measure IL-6 changes with histamine and serotonin during depression (permission from Dutta et al. 2021, Biosensors)
The Daws group also used chronoamperometry to measure serotonin changes with different pro-inflammatory cytokines and drugs that impact histamine to understand their roles with depression. For instance, they studied the cytokine inducer lipopolysaccharide (LPS) and its downstream effects on SERT [81], as well as tail suspension (TST) and forced swim tests (FST) to understand changes with LPS to depression behaviors. LPS activated SERT by increasing serotonin reuptake (faster reuptake) to subtly decrease serotonin concentrations in the mouse hippocampus (Fig. 6B). With the behavior tests, LPS injections increased immobility time, which increased depression behaviors. They also investigated the effect of increased LPS on serotonin concentrations with interleukin-1 receptor (IL-1R) deficient mice and their responses by applying SB203580 (p38 MAPK inhibitor). They found SERT activation requires IL-1R and the MAPK pathway with increased LPS. Together, these data suggest that increased pro-inflammatory LPS increases SERT activity to decrease serotonin and leads to depression behaviors that are mediated by several downstream signaling cascades, including IL-1 receptors and the MAPK pathway. Altogether, the measurements of serotonin here revealed that histamine and cytokines are critical to the molecular mechanisms of depression and are possible targets for pharmacological therapies.
Likewise, the Daws group also collaborated with several groups to compare SERT reuptake regulation by histamine receptors [82]. They used in vivo chronoamperometry in several regions of the rat brain to compare SERT activity with an H3/H4 receptor agonist (immepip) and an inverse agonist (thioperamide) (Fig. 6C). In the central nervous system, H3 receptors act as heteroreceptors to serotonin and regulate its release, but their effect on reuptake was unknown. H3/H4 receptor agonism slowed serotonin reuptake by SERT in the hippocampus, while H3/H4 receptor inverse agonism decreased serotonin concentration and caused faster serotonin reuptake by SERT [82]. The H3/H4 agonist immepip affected SERT activity only in the hippocampus and cortex, which were regulated by the CaMKII/calcineurin pathways. Altogether, the Daws and Hashemi data are similar and showed increased histamine due to inflammation, which decreases serotonin concentrations. Ultimately, bioanalytical techniques that directly measure neurochemicals show histamine is an important neuromodulator in depression, and has profound impacts on inflammation and immune cell signaling that directly change serotonin regulation through SERT activity.
Along with this work, electrochemical biosensors have also recently become popular to directly measure cytokines, which are important in inflammation [14, 15, 83]. There are several classes of cytokines, including growth factors, interferons, lymphokines, monokines, tumor necrosis factors (TNFs), and chemokines that are described in Table 1 [83], and each plays different roles in regulating cellular responses for inflammation and/or infection. In addition to depression, consistent upregulation of pro-inflammatory cytokines is associated with other serious illnesses, such as Alzheimer’s disease, cardiovascular disease, Rheumatoid arthritis, cancer, sepsis, and COVID-19 [15, 84]. Currently, many labs are designing novel biosensors to measure how cytokines change in different biofluids (saliva, serum, urine, sweat, etc.), which will aid in future experiments to explore these as biomarkers of diseases and their treatments [84]. For example, the Panchapakesan lab created and characterized a label-free IL-6 aptamer-based FET biosensor using a carbon nanotube micro-array, which is shown in Fig. 6D [85]. The biosensor consisted of a SWCNT/SiO2/Si gate coated in RNA aptamers, and IL-6 interacted with the aptamer to create a conductance that allowed IL-6 to be quantified in real-time from 1 pg/mL to 10 ng/mL. Another study, from the Zhu lab, also developed a heated carbon paste electrode with an antibody immunoassay to detect IL-6 [
留言 (0)