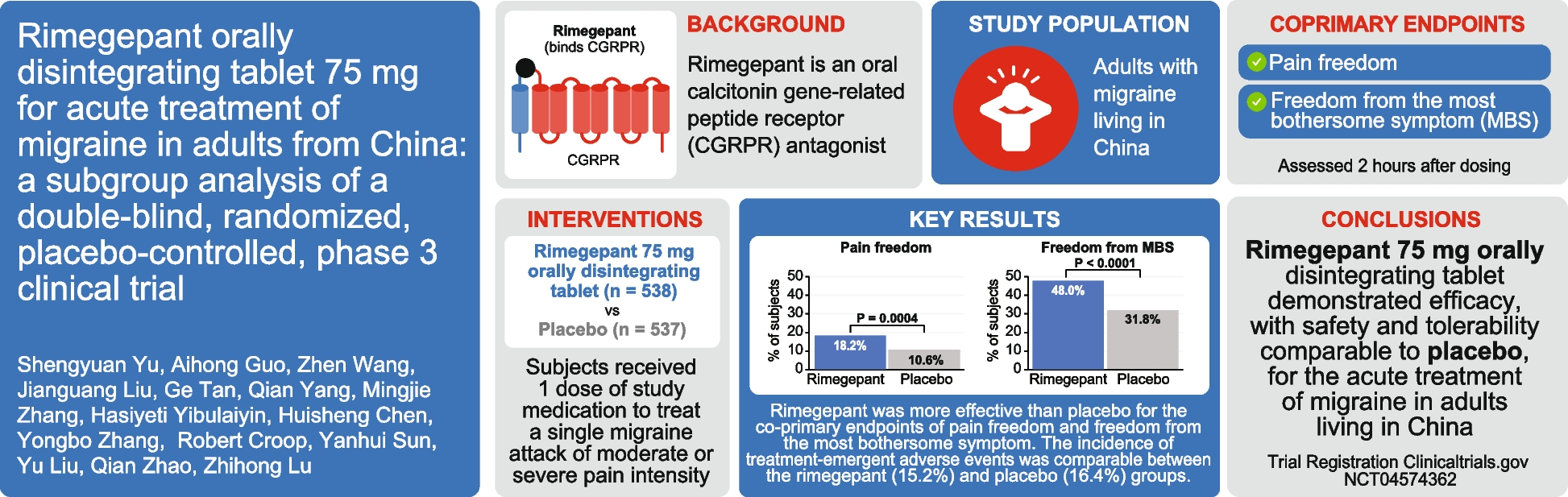
CGRP and amylin are both expressed in C1/2 DRG
This study identified the presence of amylin mRNA and peptide in C1/2 DRG neurons. Amylin-LI exhibited a puncta-like appearance in neuronal cell bodies and a pearl-like appearance in neuronal fibres, suggesting expression in vesicles, neuronal soma and fibres. Based on colocalisation with NF200 and neuronal size, amylin appeared to be present in the cell bodies of C-fibre and A-fibre neurons [8, 36]. This is the first report of amylin and CGRP co-expression in C1/2 DRG neurons, and amylin protein expression in the DRG of humans. Amylin expression in sensory ganglia has been difficult to define, in part due to anti-amylin antibodies frequently detecting CGRP at concentrations estimated to be present in neuronal vesicles [20, 21, 24,25,26,27,28,29, 39]. Hence, previously reported amylin-LI could represent false positive amylin expression at sites of high CGRP expression [16, 17, 35]. We therefore employed an anti-amylin antibody with limited CGRP cross-reactivity [28, 35]. Furthermore, amylin- and CGRP-LI did not completely overlap, with distinct immunoreactivity in discrete neurons, vesicle-like puncta, and fibres, suggesting true amylin peptide expression. However, higher exposures were required for imaging amylin compared to CGRP, suggesting amylin expression is likely modest, especially when compared to abundant neuropeptides like CGRP. RNA-FISH revealed amylin (Iapp) mRNA in rat DRG neurons. These mRNA data do not provide relative levels or the precise location of amylin peptide expression but do suggest that amylin synthesis occurs in the rat DRG [40]. Overall, multiple lines of evidence and consistency between three species indicate that amylin is expressed in upper cervical DRG, where it could play a role in transmitting sensory information, similar to CGRP. This potential physiological role is supported by the use of healthy rodents, rather than disease models.
Amylin and CGRP in DRG may signal through distinct molecular mechanisms
In addition to amylin and CGRP we report, for the first time, CTR expression in the upper cervical DRG. CTR-LI was present in the DRG of all tested species, using all tested antibodies. There were differences in the proportions of CTR-positive neuronal subtypes between species. In rodents, CTR-LI was predominantly localised in small to medium-sized neuronal cell bodies with low NF200 levels, indicating expression largely in C-fibre neurons [8, 36]. In contrast, neurons exhibiting CTR-LI in human DRG showed greater size variation, suggesting expression in both A- and C-fibre neurons, which is consistent with human single-cell DRG transcriptomics data [41, 42].
We identified that DRG neurons immunopositive for CTR were often (66–84%, Table 3) positive for amylin and/or CGRP. This could have several mechanistic implications. Firstly, both peptides may act as local agonists for CTR-based receptors in the DRG. Secondly, the presence of both ligand and receptor subunit together suggests that CGRP and/or amylin could act via autocrine mechanisms. Few studies have examined the relative distribution of the calcitonin receptor-like receptor (CLR; a component of the “canonical” CGRP receptor which is potently activated by CGRP and more weakly activated by amylin, depending on receptor species) and CGRP in the DRG. However, these studies indicate some co-expression, suggesting that CGRP may also act in an autocrine and paracrine manner at “canonical” CGRP receptors [42, 43]. Therefore, CGRP could mediate biological activity through multiple receptors and signalling mechanisms, whereas amylin could act through CTR-based receptors in a more limited population of neurons.
There are aspects of CGRP biology in sensory neurons and pain that have been difficult to fully explain by the “canonical” CGRP receptor alone. For example, autocrine autoregulatory upregulation of CGRP expression and signalling, a key feature of neuronal sensitisation and pain chronification, is only partially attenuated by CGRP receptor antagonists [9, 11]. Furthermore, CGRP promotes neuronal hyperexcitability and cortical spreading depression [44, 45]. However, fremanezumab (anti-CGRP mAb) and atogepant (CGRP receptor antagonist) cannot effectively inhibit C-fibre neuron activation [45, 46]. This suggests that CGRP could mediate some of its effects, including upregulation of CGRP and the activation and sensitisation of C-fibre neurons, through other receptors, such as the AMY receptors, in addition to the “canonical” CGRP receptor [11, 45, 46]. This aligns with our study, where amylin, CGRP and CTR appeared to be co-expressed in neurons whose size and NF200 expression suggest the C-fibre subtype in rats.
DRG and TG differentially express CGRP and amylin
The upper cervical DRG and TG innervate the head and have functional and morphological overlap as key sites for mediating craniofacial pain. However, this study, together with previous work, identifies differences in the expression of pain-related neuropeptides between these sensory ganglia [14, 17]. A notable distinction is our observation of higher abundance of amylin in the DRG, compared to the TG. Although human immunofluorescence for TG and DRG was not performed in parallel, we used the same methodology and some matched human cases for TG and DRG, suggesting differences between these cases could be genuine. There were some methodological differences for the rodent data, however, transcriptome data corroborates this difference, ranking amylin as the 8th most differentially expressed gene between rat DRG and TG [47]. Variation in amylin expression between sensory ganglia may not be unexpected as reports suggest differences in amylin expression across DRG levels [20, 21, 24]. Therefore, CTR-based receptors could be activated by two distinct ligands (CGRP and amylin) in the DRG to contribute to pain transmission, in contrast to the TG where only CGRP may be expressed, at least in the absence of disease [14, 15, 17]. Any regulation of amylin has not yet been investigated.
We observed amylin and CGRP immunoreactivity in discrete puncta, indicating different vesicle populations. This suggests there are diverse molecular processes between the DRG and TG involved in transmitting nociceptive information, such as distinct bursts of CGRP and amylin release and/or unique patterns of receptor activation. In addition, the transcription, translation, packaging, and release of amylin and CGRP could be induced under different regulatory mechanisms or temporally controlled, possibly contributing to biologically diverse aspects of pain between these sensory ganglia [48]. The expression of neuropeptides, including CGRP, in dense core vesicles is well characterised, and amylin may be present in a different subset to CGRP [49]. However, other peptides, including the related adrenomedullin, have been reported in clear synaptic vesicles in DRG fibres, which may also be the case for amylin [50]. Further study is needed to identify which vesicle subtypes contain amylin and under which conditions they are formed, regulated, and released.
Implications for treatment and future directions
Upper cervical DRG nerves, such as the greater occipital nerve, are involved in several craniofacial pain conditions, including occipital and post-traumatic brain injury-associated headaches, and migraine that predominantly affects the back of the head [1, 4, 51, 52]. These sensory neurons are involved in the pain aspects of these disorders, and other symptoms such as aura and neck stiffness [3, 51, 52]. Blocking CGRP alone may not be sufficient for craniofacial pain conditions where the DRG make a substantial contribution because DRG neurons may also express amylin and AMY receptors [3]. For example, migraine pain at the back of the head was four-fold more prevalent in response to the amylin analogue pramlintide than CGRP in a human provocation study, underscoring the potential involvement of AMY receptors in DRG-mediated pain [17]. However, it also possible that amylin and CTR-based receptors may play an anti-nociceptive role because exogenous amylin reduced nociceptive behaviour when administered prior to noxious stimuli, such as formalin and acetic acid [19, 22, 23]. In addition, administration of CTR-based receptor agonists, such as salmon calcitonin, have analgesic properties and are reported to decrease TG and DRG neuron activation [53,54,55,56,57]. This suggests that the contributions that amylin and the CTR-based receptors make to pain and migraine are likely complex and further research into this system is warranted.
Determining which receptors are present and by what mechanisms they contribute to nociceptive signalling and sensitisation is crucial for developing therapeutics. Our signalling data indicate expression of functional CGRP- and amylin-responsive receptors in the DRG. The pharmacology suggests functional CTR expression, potentially as part of AMY receptors, as amylin has relatively limited activity at the rodent CGRP and adrenomedullin receptors, while both CGRP and amylin are equipotent at some rodent AMY receptors [37, 38]. However, the pharmacology of these receptors is complex, and there are some species differences meaning that it is difficult to draw firm conclusions about the role of individual receptors. CLR mRNA and protein has been detected in DRG neurons, therefore, CGRP could potentially signal through CLR and/or CTR-based receptors [42, 43]. Examining RAMP expression in conjunction with CTR and CLR subunits to determine the spatial distribution of receptor-RAMP pairs could help reveal which receptors are relevant. However, many anti-RAMP and anti-CLR antibodies have limitations [33, 58, 59]. The absence of CLR and RAMP immunofluorescence in this study prevents further delineation of the role of each receptor in CGRP and amylin mediated signalling in the DRG. Future studies could consider non-antibody-based methods, such as fluorescent in situ hybridization, mass spectrometry imaging and spatial transcriptomics or proteomics. In addition, the potential for paracrine and autocrine signalling mechanisms should be considered as autocrine signalling is proposed to require higher antagonist concentrations to attenuate receptor activation, relative to paracrine signalling [60].
Our study noted some species and sex differences. For example, we observed a greater co-expression of CTR with amylin, and of peptides in C-fibre neurons in female rats, which may underlie the sex-dependent differences in amylin sensitivity previously reported [17, 18]. We did not use pain models or human cases with a migraine diagnosis. However, previous studies have reported upregulation of CGRP and amylin in the DRG in response to noxious stimuli [20, 21]. In addition, sex-specific and pain-specific differences in expression of the CGRP and amylin peptide and receptor systems in human DRG have been reported [42]. Another consideration is the age of the human cases examined, as the expression of neuropeptides is known to change during ageing [61, 62]. Examination of the spatial relationships of this family of peptides and receptors in younger, and migraine patients will shed additional light on their contribution to nociceptive signalling and potential as therapeutic targets.





留言 (0)