
記住我
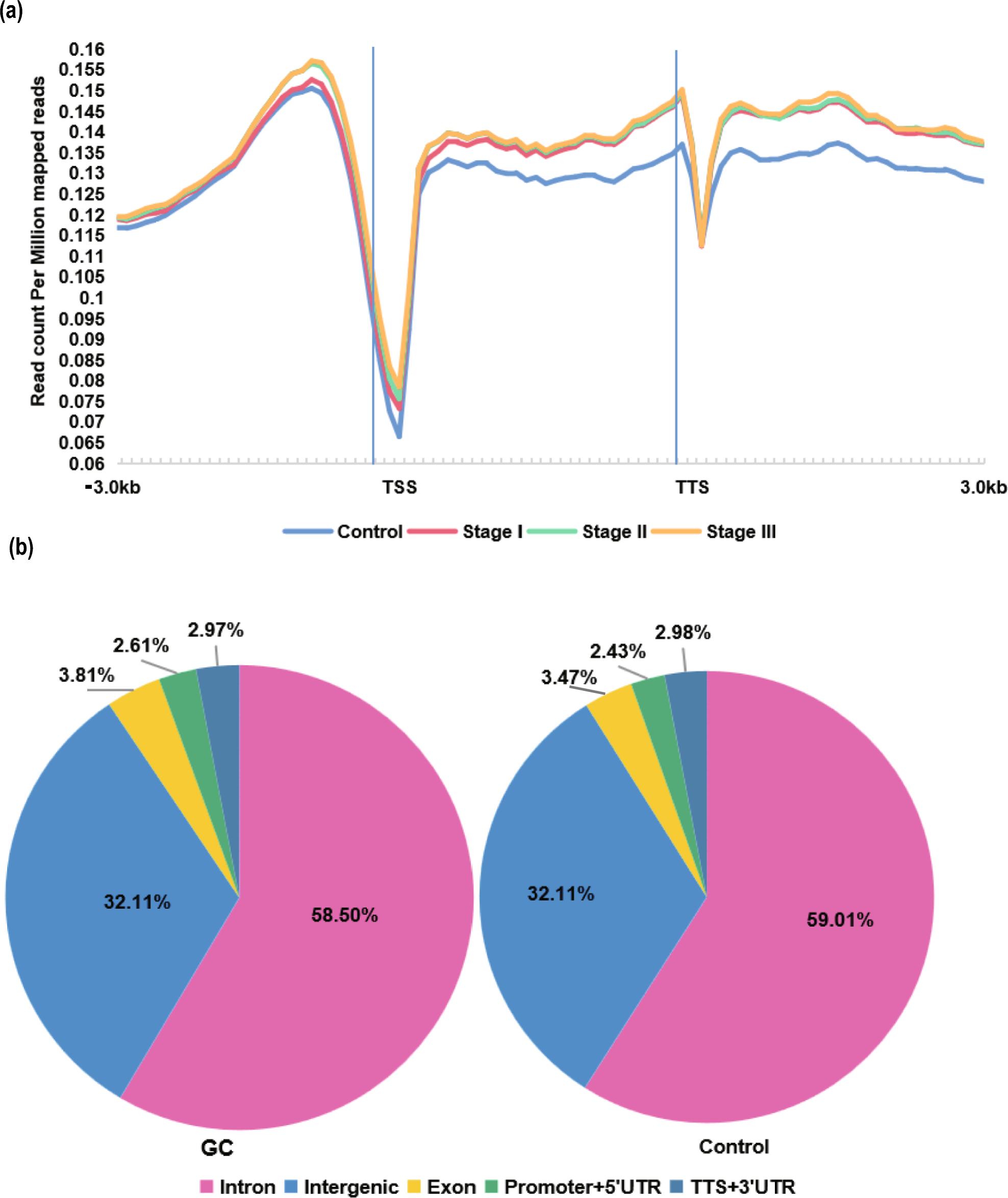


Previous bulk transcriptional sequencing (RNA-seq) identified the overexpressed immune checkpoints TIM-3 and VISTA in ascitic cells of patients with PM-GC, indicating ascitic cells’ high immune escape ability [9]. To re-confirm this, aDTCs from ascites and CTCs from paired PB samples of 33 treatment-naïve patients with PM-GC were enriched via the previously established SE-iFISH platform (SE-iFISH cohort, Supplementary Fig. S1 and Table S2). Additionally, a pairwise comparison of the expression of PD-L1, another immune checkpoint marker mediating immune escape [22, 23], was conducted on enriched aDTCs or CTCs. The enumeration total and expression levels of PD-L1+ on aDTCs or ascites-disseminated microemboli (aDTMs), also known as ascites-disseminated tumor cell clusters aggregated with two or more aDTCs, were all higher than those of CTCs or circulating tumor microemboli (CTMs) [24] (Fig. 1a–d). In individual patients, the expression level of PD-L1 on aDTCs and aDTMs were augmented compared with that on paired CTCs and CTMs (Fig. 1e).
Fig. 1
Upregulation of PD-L1 on aDTCs mediates immune escape and deteriorating prognosis in PM-GC. a and b Quantitative comparison of aDTCs (a) and aDTMs (b) in ascites (N = 33) with CTCs (a) or CTMs (b) in the paired peripheral blood samples (N = 33). Data was statistically analyzed using the non-parametric Mann–Whitney test. ***P < 0.001. c and d Comparison of proportions of PD-L1 + aDTCs (c) or aDTMs (d) within total aDTCs or aDTMs from ascites samples (N = 33) to PD-L1 + CTCs (c) or CTMs (d) within total CTCs or CTMs from paired peripheral blood samples (N = 33). e Representative immunofluorescent images showing PD-L1 expression on aDTCs or aDTM and CTCs or CTM from the same patient using SE-iFISH (bar = 5 µm). f Proportions of chromosome 8 (chr 8) triploidy, tetraploidy, and multiploidy in total detected PD-L1+ aDTCs from ascites samples (N = 33, upper pie chart) or PD-L1+ CTCs from paired peripheral blood samples (N = 33, lower pie chart). g Kaplan–Meier plots illustrating the overall survival of patients with negative PD-L1+ aDTCs and that of those with positive PD-L1+ aDTCs. h Proportional variations of PD-L1+ aDTCs following treatments. (Abbreviations: aDTC ascites-disseminated tumor cells, CTS cathepsin, CTM circulating tumor microemboli; aDTC; CTC; aDTM; CTM; aDTCtri and CTCtri are chr 8 triploid aDTC and CTC; aDTCtetra and CTCtetra are chr 8 tetraploid aDTC and CTC; aDTCmulti and CTCmulti are chr 8 multiploid aDTCs and CTCs.)
Besides enumeration and expression levels, the karyotypic characteristics of PD-L1+ on aDTCs and CTCs are completely distinct. PD-L1+ CTCs are predominantly multiploid chromosome 8 (chr8) (CTCmulti), whereas PD-L1+ aDTCs are mainly triploid chr8 (aDTCtri), indicating an elevated therapeutic tolerance of PD-L1+ aDTCs (Fig. 1f) [25]. Moreover, the prognostic analysis of the SE-iFISH cohort (Cohort-1) confirmed shorter overall survival (OS) of patients with positive pre-therapeutic PD-L1+ aDTCs (PD-L1+ aDTCs ≥ 1 cell/6 mL, Fig. 1g). These results are in line with previous findings and further demonstrated that the overexpressed PD-L1 on aDTCs is involved in aDTC immune evasion.
Current studies on PD-L1+ tumors suggest a paradoxical role of the PD-L1 expression on the PD-1/PD-L1 blockade efficiency [26, 27]. Although higher pre-therapeutic PD-L1 levels in primary tumors are considered an indicator of increased sensitivity to anti-PD-1/PD-L1 therapy, the re-acquisition or uncontrollable PD-L1 expression on either tumor or immune cells is associated with therapeutic resistance to PD-1/PD-L1 blockades [27]. Our results indicated that disruption of tumor-cell intrinsic aggressiveness by systematic chemotherapies could not abolish overexpressed PD-L1 on aDTCs, thereby demonstrating that PD-L1+ aDTCs were still detectable in most post-therapy ascites in 5 of 33 enrolled patients (Fig. 1h). This implies that re-acquisition or uncontrolled PD-L1 overexpression may be regulated by tumor cell-intrinsic mechanisms and tumor cell-extrinsic mechanisms. Therefore, inhibiting only tumor-cell intrinsic targets cannot completely curb aDTC immune escape.
MPs dominate the TME of aDTC-rich ascitesTo comprehensively understand tumor-cell extrinsic mechanisms underlying aDTC immune evasion, we cataloged the cell components of ascites and paired PB samples collected from seven treatment-naïve patients with PM-GC (scRNA-seq test cohort, Supplementary Fig. S1 and Supplementary Table S3) by leveraging single-cell transcriptional sequencing (scRNA-seq). Overall, 144,522 cells from all 14 ascites samples and matched PB samples, which passed multiple quality controls, were further clustered using Seurat. Overall, eight cell clusters, including MPs [28], epithelial cells [29], T cells [30], B cells [31], platelets [32], fibroblasts [33], erythrocytes [32], and neutrophils [34], were annotated using canonical markers (Fig. 2a, Supplementary Fig. S3A, and Supplementary Table S1). Among these, epithelial cells and MPs were mainly from ascites samples, while T and B cells were from both ascites and PB samples (Fig. 2b, Supplementary Fig. S3B-C).
Fig. 2
scRNA-seq of seven treatment-naïve ascites and paired peripheral blood samples from the scRNA-seq test cohort. a and b UMAP representation of all annotated cell clusters from seven ascites and paired peripheral blood samples. Cell clusters are colored according to cell type (a) or sample origin (b). c Unsupervised clustering of epithelial cells from ascites. The upper UMAP is colored according to epithelial cell clusters, and the lower UMAP is colored according to different ascites samples. d Copy number variations (CNV) in epithelial cell clusters. Columns represent epithelial clusters, and rows represent chromosomal regions. The CNVs of CD45+ cells serve as references. e The proportion of PD-L1+ aDTCs in total aDTCs detected in seven treatment-naïve ascites from the scRNA-seq test cohort using SE-iFISH. Each column represents a unique ascites sample, and the two rows represent the proportion of PD-L1+ aDTCs and PD-L1− aDTCs, respectively. f Histograms displaying the proportions of aDTCs, T cells, and mononuclear phagocytes (MPs) in seven ascites samples from the scRNA-seq test cohort. The absolute number of aDTCs, T cells, and MPs is shown in Supplemental Fig. 2C. (Abbreviations: aDTC ascites-disseminated tumor cells, ASSubtype1 aDTC-rich ascites, ASSubtype2 MP-dominant ascites, ASSubtype3 T-dominant ascites, UMAP Uniform Manifold Approximation and Projection)
Since aDTC presence is a prognosticator of an inferior OS in PM-GC (Supplementary Fig. S3D), we first inspected the malignant subsets in epithelial cells. In the predominantly enriched samples AS_1, AS_4, and AS_2, 16 epithelial cell clusters were identified (Fig. 2c and Supplementary Fig. S3C). Consistent with previous findings [10], the epithelial cells in the ascites samples from PM-GC patients were highly heterogenous, and each ascites sample comprised specific epithelial cell clusters. For instance, the epithelial cell clusters E2, E3, E4, and E9 were only found in sample AS_1. Conversely, the clusters E6, E10, and E14 belonged exclusively to sample AS_4 (Fig. 2c and Supplementary Fig. S3E). High diversity of CNVs was identified in the individual clusters, further demonstrating malignancy in all 16 epithelial cell clusters; accordingly, we referred to all epithelial cell clusters as aDTC clusters (Fig. 2d). The 17q copy number gain, which is considered a unique event in tumor cells from the stomach [10], was identified in most aDTC clusters in our study, confirming the stomach tumor-specific origin of all detected aDTC clusters (Fig. 2d).
Regarding PD-L1 expression, we failed to recognize the transcriptional PD-L1 elevation in aDTC clusters, probably owing to the dropouts in scRNA-seq. However, PD-L1 protein overexpression on aDTCs was validated by SE-iFISH, as demonstrated by > 50% of aDTCs being detected as PD-L1-positive in five out of seven ascites samples (Fig. 2e).
We hypothesized that the upregulated PD-L1 on aDTCs and its immune escape mediation is modulated by immunosuppressive interactions between the cell-extrinsic immune TME and aDTCs. To demonstrate this, the immune components, especially in the aDTC-rich ascites samples (samples AS_1, AS_4, and AS_2: referred to as ASSubtype 1), were analyzed. MPs, rather than T cells, were overwhelmingly infiltrated in the TME of the aDTC-rich ascites samples (Fig. 2f and Supplementary Fig. S3C). Moreover, aDTC quantification in aDTC-rich ascites showed an inverse correlation with the presence of MPs, implying robust communications between aDTCs and MPs (Fig. 2f).
In aDTC-deficient ascites, MPs and T cells may exist exclusively in the TME of the ascitic fluid, which further shapes the two heterogenous TME subtypes of ascites: ASSubtype 2 and ASSubtype 3. In ASSubtype 2 ascites, the TME is MP-dominant, with < 10% T cells being infiltrated (sample AS_3, AS_6, and AS_5, Fig. 2f). In ASSubtype 3 ascites, a high presence of T cells in the TME impedes MP infiltration (sample AS_7, Fig. 2f). These results indicated that MPs and T cells might independently modulate the development of ascites in patients with PM-GC.
Ascitic TAMs form a continuum transitioning from cathepsin (CTS)high to complement 1q (C1Q) highPhysiologically, a macrophage plays a critical role in maintaining immune homeostasis in the peritoneal cavity [1]. This raises the question of whether TAMs are hub MPs involved in immunosuppressive communication with aDTCs in PM-GC ascites. Moreover, macrophages account for the highest MP proportion, especially in the aDTC-rich ASSubtype 1 ascites (Fig. 3a, b and Supplementary Fig. S4A). Additionally, the upregulated DEGs identified in macrophages, including APOE, C1QB, APOC1, C1QC, C1QA, SPP1, LYVE1, LGMN, and cathepsin L (CTSL), are all anti-inflammatory, further conferring their immunosuppressive function; accordingly, we referred to all annotated macrophages as TAMs (Fig. 3c). To disclose the specific function of the individual TAM lineage clustered by unsupervised learning (Fig. 3d and Supplementary Fig. S4B), we initially examined canonical M1 and M2 signatures in each TAM cluster [16]. Nevertheless, all TAM clusters failed to fall into the M1/M2 dichotomy (Supplementary Fig. S4C and Supplementary Table S4). However, when comparing DEGs of TAMs from the MP clustering (Fig. 3c) in the individual TAM lineage, we noted the presence of transitionally varied expression patterns of the C1Q-associated genes C1QA, C1QB, and C1QC and CTS-associated genes CTSA, CTSD, CTSL, which can confer three TAM lineages perfectly: CTShigh/C1Qhigh (Macro 6), CTSlow/C1Qhigh (Macro 3 and Macro 2), and CTShigh/C1Qlow (Macro 1, Macro 4, and Macro 5) (Fig. 3e). These indicated that the expressional variation of CTS and C1Q genes on ascitic TAMs are a continuum instead of a dichotomic polarization.
Fig. 3
scRNA-seq analysis of the ascitic TAMs of seven treatment-naïve ascites from the scRNA-seq test cohort. a UMAP representation of annotated subpopulations of MPs. Different colors are correlated to different cell types of MPs. b Proportions of individual MP subpopulations in three ascitic subtypes. c Heatmap showing the top 10 differentially expressed genes (DEGs) in each MP subpopulation. Columns represent normalized expressions of DEGs, and each row represents a cell. Mono, monocytes; Macro, macrophage; cDC2, type 2 conventional dendritic cell; mDCs, mature dendritic cells. d The UMAP of the unsupervised clustering of macrophages. Each color represents a unique macrophage cluster. e Average expressions of DEGs in macrophages identified from panel c in the individual macrophage cluster. Each column represents the average gene expression normalized by Z score normalization, and each row represents a unique macrophage cluster. f Trajectory analysis of the differentiation of TAMs. Cells on the trajectories are colored according to their states (outer graph) or pseudotime (inner graph). g Respective projection of C1Qhigh/CTShigh (Macro 6), C1Qhigh (Macro 2 and 3), and CTShigh (Macro 1, 4, and 5) TAM lineages onto the trajectory of TAM differentiation. h Variation of the expressions of C1Q or CTS genes following pseudotime. Each dot in the graphs represents a unique cell. Color coding is explained in (g). (Abbreviations: MPs mononuclear phagocytes, TAM Tumor-associated macrophages, UMAP Uniform Manifold Approximation and Projection
Therefore, we investigated how this continuum of gene expression varied following TAM lineage differentiation. The trajectory of TAM lineages analyzed using Monocle 2 is shown in Fig. 3f. The five developmental states of TAM lineages were determined. The differentiation of ascitic TAMs commenced from the CTShigh/C1Qhigh Macro 6, evolving to CTShigh TAM lineages (Macro 1, 4, and 5) at intermediate states (Fig. 3g and Supplementary Fig. S4D). Distinct from the CTShigh TAM lineages that exist in all states thereafter, the C1Qhigh TAM lineages (Macro 2, and 3) were only developed in the terminal state (Fig. 3g and Supplementary Fig. S4D). Further insight into the expression of C1Q and CTS genes following pseudotime confirmed that C1Q gene levels were only elevated at the terminal pseudotime. This was contrary to the CTS genes, which showed a terminal decrease after an increase at the intermediate pseudotime (Fig. 3h).
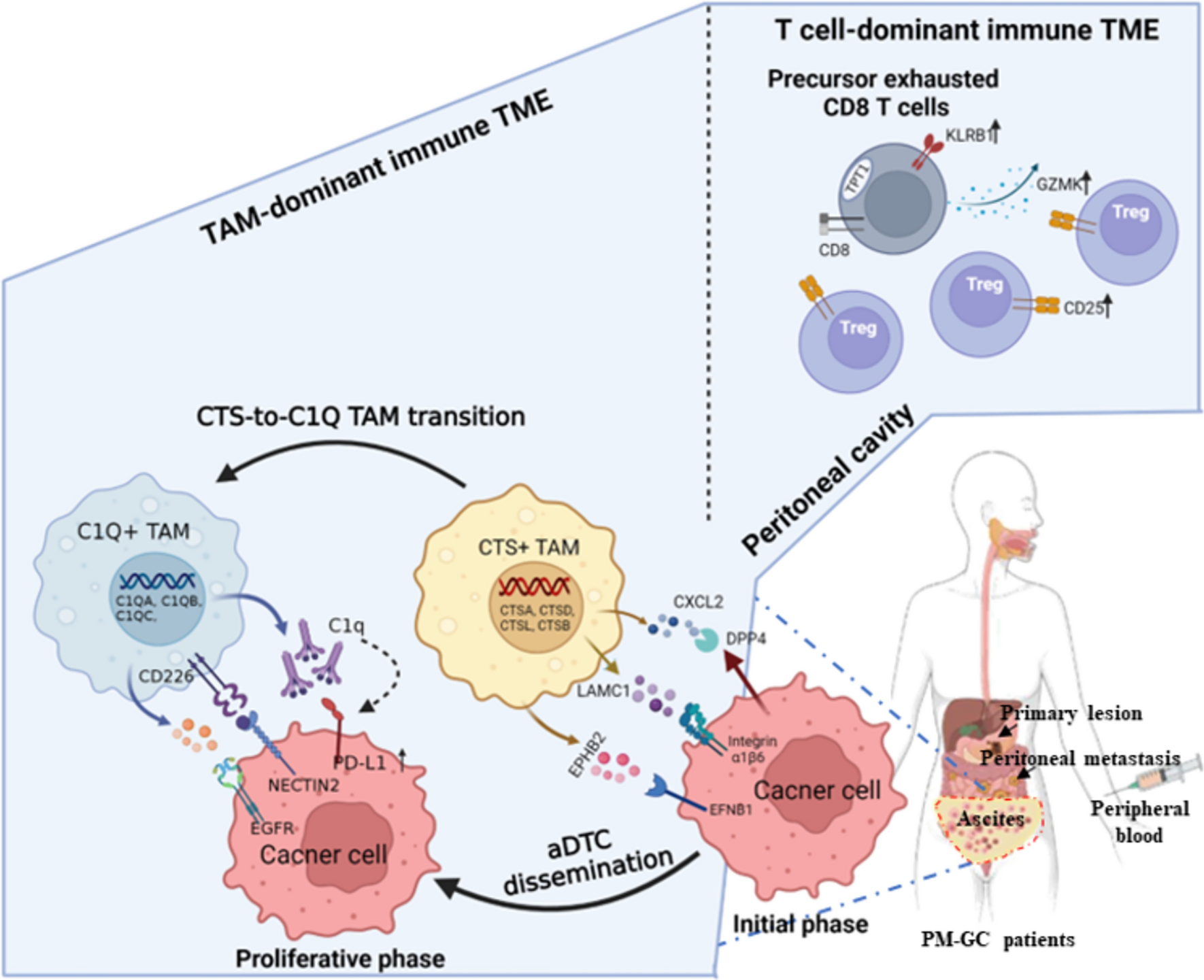
Altogether, the obtained results support the view that ascitic TAMs undergo a lineage transition from the CTShigh to C1Qhigh subtype (CTS-to-C1Q transition) and that the CTShigh lineages are TAM precursors, which differentiate terminally toward the C1Qhigh lineages.
TAM transition from CTShigh to C1Qhigh modulates different aDTC dissemination phasesHaving uncovered the specific CTS-to-C1Q transition in ascitic TAMs, we next investigated how the TAMs in different transitional states interact with aDTCs and influence their dissemination. To this end, the cell-to-cell communications (CCC) between TAMs and aDTCs were analyzed using the Cell-PhoneDB in an aDTC-rich AS Subtype 1 ascites. The interaction strength between TAM lineages and aDTCs is mapped in Fig. 4a and Supplementary Fig. S4E, which show that the interaction strength of aDTCs with CTShigh (Macro 1) and C1Qhigh (Macro 2) lineages is comparable (Fig. 4a). Similarly, a comparison of the interaction types and directionality between the CTShigh or C1Qhigh lineages and aDTCs did not show any difference (Fig. 4b and Supplementary Fig. S4F). These results inferred that CTShigh and C1Qhigh TAMs crosstalk robustly with aDTCs. CTShigh or C1Qhigh TAMs are more likely the signal-sending cells propagating secreted signaling to modulate aDTC behavior.
Fig. 4
C1Qhigh TAMs mediate the upregulation of PD-L1 and NECTIN2 on aDTCs. a Chord diagrams representing cell–cell communication (CCC) between aDTCs and TAM clusters identified in Fig. 3e in individual ASSubtype 1 ascites. Different colors on the outer circle of the chord diagram are correlated to different cell clusters. Chords curved inside the circle represent interactions between two cell clusters they connected. Wider chords mean stronger interactions. b Donut plots displaying proportions of secreted signaling and non-secreted signaling from aDTCs to TAMs (aDTCs (R) > TAMs (L)) or vice versa (TAMs (R) > aDTCs (L)) in each ASSubtype 1 ascites sample. c Top 30 interaction pairs between C1Qhigh (labeled as C1Q in the graph) or CTShigh (CTS) TAMs and aDTCs (Labeled as E). d Specific interaction pairs between C1Qhigh (C1Q) or CTShigh (CTS) TAMs and aDTCs (Labeled as C1Q in the graph) or CTShigh (CTS) TAMs and aDTCs (E). In C and D, the sending cells and signals are labeled red. e Expressional correlations of C1Q or CTS genes to PD-L1 in 48 metastatic GC from TCGA database. Red means a positive correlation, and blue represents a negative correlation. Data are correlated using Pearson’s correlation coefficient. (*P < 0.05; **P < 0.01; ***P < 0.001). f Representative imaging of in vitro cultured aDTCs. Left panels: The morphology of in vitro cultured aDTCs. Right panels: Identification of in vitro cultured aDTCs using SE-iFISH. The bars in the left panels are 200 µm, and those in the right panels are 5 µm. g Protein levels of PD-L1 and NECTIN2 on 4 in vitro cultured aDTCs with or without treatments of corresponding ascites supernatants (Ascites #1, #2, #3, and #4) detected by western blot. h and i Quantification of the band intensities of PD-L1 and NECTIN2 from (g). The band intensities are determined by the software ImageJ with GAPDH as the standard (Mean ± SD, N = 3, paired t—test was used for statistical analysis, *P < 0.05; **P < 0.01; ***P < 0.001; N = 3). (Abbreviations: aDTC ascites-disseminated tumor cells, CTS cathepsin, MPs mononuclear phagocytes, TAM tumor-associated macrophages, UMAP Uniform Manifold Approximation and Projection)
Accordingly, we examined the secreted crosstalk between CTShigh or C1Qhigh TAMs and aDTCs. Figure 4c, d displays the top 30 difference or specifically interaction pairs between TAM lineages and aDTCs. CTShigh and C1Qhigh TAMs generate immunosuppressive interactions with aDTCs via predominantly secretion of SPP1 [35], FN1 [36], TIMP1 [37], or upregulation of CD74 [38], all of which foster tumor-cell metastasis (Fig. 4c). Specifically, CTShigh TAMs are inclined to secrete CXCL2 [39], LAMC1 [40], and NRP1 [41], which shape the TME of pre-metastatic niches and facilitate angiogenesis [42, 43]. Additionally, CTShigh TAMs secrete EPHB2, which contributes crucially to tumor-cell stemness via engagement to its receptor EFNB1 [44]. C1Qhigh TAMs interact principally with aDTCs via the epiregulin-epidermal growth factor receptor (EGFR) [45] and CD226-NECTIN2 receptor–ligand pairs, which are involved in tumor-cell survival and proliferation [46, 47] (Fig. 4d).
Collectively, we suggested that CTShigh TAMs likely regulate the initial phases of aDTC dissemination via programming pre-metastatic TME and attracting aDTCs homing into ascites, while the stepwise transition from CTShigh to C1Qhigh TAMs triggers and maintains the overproliferation of homed aDTCs in ascites.
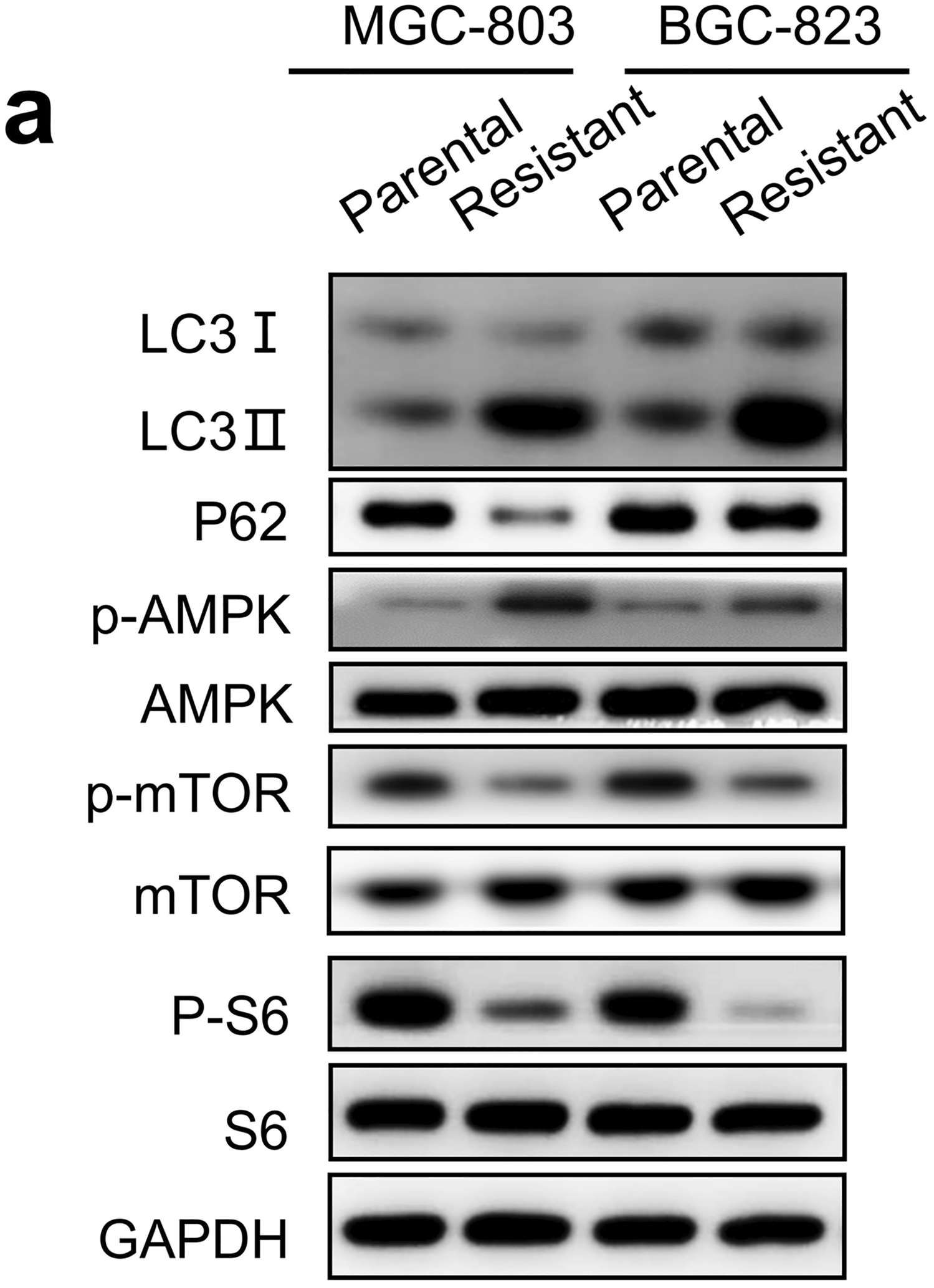
C1Qhigh TAMs drive aDTC immune evasion in the proliferative phase of ascitic metastasisDue to the crucial role of the CTS-to-C1Q transition in the TAM-aDTC crosstalk, we next investigated whether this transition was also involved in the aDTC PD-L1-mediated immune escape, as demonstrated in the SE-iFISH cohort (Fig. 1). By analyzing RNA-seq data of 48 metastatic GCs from The Cancer Genome Atlas (TCGA) database, we first noted that C1Q genes (C1QA, C1QB, and C1QC), rather than CTS genes (CTSA, CTSB, CTSD, and CTSL), showed a positive correlation with the PD-L1 levels (Fig. 4e). To confirm further the involvement of C1Qhigh TAMs in the PD-L1 upregulation on aDTCs, we cultured aDTCs from eight ascites of PM-GC samples in vitro (Fig. 4f). Treatments of the ascitic supernatants significantly enhanced PD-L1 expression on the in vitro cultured aDTCs (Fig. 4g, h and Supplementary Fig. S4G-H). Similarly, NECTIN2, which interact specifically with C1Qhigh TAMs, expressions were upregulated in ascites-treated aDTCs (Fig. 4g, i, Supplementary Fig. S4G and S4I). This synchronous upregulation of PD-L1 and C1Qhigh TAM-interacting proteins was also observed in ascites-treated GC cell lines HGC27, MKN45, and SNU1 (Fig. 5a–c), confirming that PD-L1 upregulation on aDTCs occurs in the proliferative phase of ascitic metastasis, coincident with the C1Qhigh TAMs-driven overproliferation of aDTCs.
Fig. 5
C1q participates in PD-L1 and NECTIN2 upregulation on aDTCs. a Protein levels of PD-L1 and NECTIN2 on 3 GC cell lines (HGC27, MKN45, and SNU1) with or without treatments of ascites supernatants (Ascites #1, #3, and #4) detected by western blot. b and c Quantification of the band intensities of PD-L1 (b) and NECTIN2 (c) from (a). d Correlations between protein levels of complement proteins C1q, C2, C3, and C4 detected by ELISA and the enumeration of PD-L1+ aDTCs or aDTMs in the SE-iFISH cohort. Red means a positive correlation, and blue represents a negative correlation. Data are correlated using Pearson’s correlation coefficient. (*P < 0.05; **P < 0.01; ***P < 0.001). e Workflow diagrams of TurboID biotin labeling system for interaction proteomics. TurboID genes are transfected into in vitro cultured aDTCs with lentivirus. Overexpressed TurboID in the transfected aDTCs can biotinylate neighboring proteins in ascites supernatant interacting with aDTCs. Then, the biotinylated proteins are isolated by streptavidin-coated beads and analyzed using LC–MS/MS. f Western blots showing efficient biotinylation in HEK293T transfecting with TurboID vector compared with those with empty vector (EV). g Western blots showing biotinylated proteins enriched from in vitro cultured aDTCs treated with or without paired ascites supernatant in the presence or absence of biotin. h Histogram showing the relative abundance of C1QA, C1QB, C1QC, C3, and C4 in biotinylated proteins enriched from in vitro cultured aDTCs treated with or without pairing ascites. Levels of complement components in enriched proteins are detected using LC–MS/MS, and fold changes are used to display the relative abundance of complement proteins. (Mean ± SD, N = 4, Mann–Whitney test was used for statistical analysis. *P < 0.05). i Protein levels of PD-L1 and NECTIN2 on 3 GC cell lines (HGC27, MKN45, and SNU1) treated with or without ascites #2 after adding recombinant complement C1q cytokines. j and k Quantification of the band intensities of PD-L1 (j) and NECTIN2 (k) in (i). l Schematic diagram showing co-culturing of gastric cancer cell lines with THP-1 derived macrophages in the presence or absence of human recombinant C1 inhibitor (C1NH). m Expression of PD-L1 and NECTIN2 on MKN45 and SNU1 cell lines co-cultured with THP-1 derived macrophages with different concentrations of C1NH. n and o Quantification of the band intensities of PD-L1 (n) and NECTIN2 (o) in (m). All the bands intensities are determined by the software ImageJ with GAPDH as the standard (Mean ± SD, N = 3, two-way ANOVA test was used for statistical analysis, *P < 0.05; **P < 0.01; ***P < 0.001). (Abbreviations: aDTC ascites-disseminated tumor cells, LC–MS/MS liquid chromatography-mass spectrometry/mass spectrometry)
Complement protein C1q participates in PD-L1 upregulation on aDTCs driven by C1Qhigh TAMsNext, we investigated whether C1Qhigh TAMs modulate PD-L1 and NECTIN2 overexpression on aDTCs by secreting complement protein C1q in the SE-iFISH cohort. The levels of C1q and C3 in ascites both showed a positive correlation with the number of PD-L1+ aDTCs and aDTMs (Fig. 5d), suggesting that the activation of the appropriate classical complement pathway underlies PD-L1 upregulation on aDTCs.
To further illustrate the direct interaction of C1q and ascitic tumor cells, the biotin ligase TurboID labeling system [28] was transfected into in vitro cultured aDTCs (Fig. 5e). The overexpressed TurboID biotinylates neighboring cytokines in ascites supernatant that interacts with aDTCs. Further proteomic analysis of the biotinylated cytokines enriched by streptavidin magnetic beads (Fig. 5f, g) showed that complement proteins C1QA, C1QB, C1QC, C3, and C4 were all enriched in ascites-treated cells (Fig. 5h), demonstrating the direct crosstalk between components in the complement pathway and ascitic tumor cells.
The next question is whether the engagement of complement proteins influences PD-L1 and NECTIN2 expression on ascitic tumor cells. To address this, we compared PD-L1 and NECTIN2 expressions on GC cell lines treated with either singular ascitic supernatants or their combination with the recombinant C1q protein. Adding recombinant complement C1q protein to GC cell lines HGC27, MKN45, and SNU1 treated by ascites #2 can further augment PD-L1 and NECTIN2 upregulation induced by singular ascitic supernatants (Fig. 5i–k).
To further confirm the role of C1Qhigh TAMs, GC cell line MKN45 and SNU1 were co-cultured with THP-1 derived macrophages in the presence or absence of C1NH, the human recombinant C1 inhibitor used to abolish the function of secreted C1q from macrophages (Fig. 5l). As shown in Fig. 5m–o, addition of C1NH significantly mitigates both PD-L1 and NECTIN2 expression on MKN45 and SNU1, which further confirms that C1q secreted by co-cultured macrophages mediates the activation of complement pathway, participating in the PD-L1 and NECTIN2 upregulation and promoting aDTCs immune evasion in PM-GC.
CTS-to-C1Q transition of TAMs fosters therapeutic resistance of PM-GCHaving determined the contribution of the CTS-to-C1Q transition of TAMs to aDTC dissemination and immune escape, we next investigated whether this transition could lead to therapeutic resistance in patients with PM-GC. To validate the CTS-to-C1Q transition of ascitic TAMs specifically, we used another independent cohort containing both treatment-naïve and paired therapeutically resistant ascites samples from eight patients with PM-GC (scRNA-seq validation cohort, Supplementary Fig. S1 and Supplementary Table S5). Figure 6a displays TAM lineages clustered from 18 ascites samples from the scRNA-seq validation cohort. Comparison of TAM lineages from treatment-naïve or therapeutically resistant ascites patients highlights cluster Macro 4 as the only therapeutically resistant (TR) TAM lineage, which was determined by its proportional elevation in the therapeutically resistant ascites (Fig. 6b, c and Supplementary Fig. S5A). Transcriptional levels of C1QA, C1QB, and C1QC showed a marked increase in Macro 4 (Fig. 6d, e). Conversely, neither of the CTS genes showed variations in Macro 4 compared with the other TAM lineages (Fig. 6e).
Fig. 6
scRNA-seq analysis of the TAMs from the treatment-naïve and paired therapeutically resistant ascites of eight patients with PM-GC in the scRNA-seq validation cohort. a and b UMAP representation of unsupervised clustered macrophage lineages from all treatment-naïve and therapeutically resistant ascites. Different colors in (a) are correlated to different macrophage lineages, and those in (b) represent pre-therapeutic or resistant (progressive disease, PD) ascites. c Proportional variations of each TAM cluster after therapeutic resistance, categorizing TAM clusters into three subtypes: conservative TAMs (the proportions are almost unvaried after therapeutic resistance), therapeutically sensitive (TS) TAMs (clusters that can be reduced after therapy), and therapeutically resistant (TR) TAMs (the proportion elevated after therapeutic resistance). d Top 10 DEGs identified in conservative, TS, and TR TAMs. All genes shown in this heatmap are listed in detail in Supplementary Table 6. Each column represents a unique cell, and each row represents the normalized expression of each gene. e Expression levels of CTS and C1Q genes in conservative, TS, and TR TAMs. f Trajectory analysis of TAM differentiation following therapeutic resistance. Trajectories are, respectively, colored according to pseudotime (upper panel) and developmental states (lower panel). g Sankey diagram displaying the dynamic evolution of dominant TS (Macro 5, 8, 6, and 9) and TR TAMs (Macro 4) following therapeutic resistance. Nodes with different colors in the left and middle columns represent different TAM lineages and developmental states, respectively. h Variations of CTS and C1Q genes following pseudotime. (Abbreviations: CTS cathepsin, DEGs differentially expressed genes, TAM tumor-associated macrophages, TME tumor microenvironment, UMAP Uniform Manifold Approximation and Projection)
Next, we investigated whether C1Qhigh TR TAMs transitioned from the CTShigh lineages, leading to therapeutic resistance. Figure 6f shows the trajectory of the TAM differentiation following therapeutic resistance. The therapeutically sensitive (TS) Macro 5, identified by its large decrease in TR ascites level (Fig. 6c), is predominant in the TAM precursor lineage, presenting in the initial state (State 1 in Fig. 6f and Supplementary Fig. S5B-C). Without treatments, Macro 5 directly develops to another TS lineage, Macro 8, existing mainly in State 5 (Fig. 6g and Supplementary Fig. S5B-D). Other TS TAM lineages (Macro 6 and Macro 9) showed progression similar to that in State 5 (Fig. 6g). However, under therapeutic pressures, Macro 5 differentiates distinctly into the TR TAM lineage Macro 4 at the bifurcated States 2, 6, and 7 (Fig. 6f, g and Supplementary Fig. S5D). Further insight into the variations in C1Q and CTS gene levels following pseudotime shows that the CTS genes showed high expression in the initial pseudotime, when TS lineages were predominant. However, this overexpression of the CTS genes gradually transitioned to an overexpression of the C1Q genes at the intermediate pseudotime when the TR TAM lineage was developed (Fig. 6h).
Collectively, the results demonstrated that the TAM transition from the CTShigh to C1Qhigh lineages facilitates the development of therapeutic resistance and ascitic progression in patients with PM-GC.
Ascitic CD8+ T cells are dysfunctional, showing limited cytotoxic activityAs mentioned previously, the exclusive infiltration of T cells in ASSubtype 3 ascites suggests their independent function in ascites development. Therefore, we explored the specific characteristics of ascitic T cells based on our scRNA-seq data from the scRNA-seq test cohort (seven treatment-naïve ascites and paired PB samples, Supplementary Fig. S1 and Supplementary Table S3). Overall, 12 T-cell clusters were identified in the ascites and paired PB samples (Fig. 7a and Supple
留言 (0)