Human mitochondrial DNA (mtDNA) is a circular DNA molecule with 16,569 base pairs. mtDNA encodes a set of RNAs and essential protein subunits in the oxidative phosphorylation system. mtDNA is susceptible to genotoxic agents and reactive metabolites and is maintained by DNA repair, mtDNA turnover, and mitochondrial dynamics [1], [2], [3], [4], [5], [6]. The susceptibility to genotoxins, multicopy characteristics, and the signaling role of mtDNA in immune response and inflammation have led to the proposed role of mtDNA as a genotoxic stress sensor in cells [7], [8].
A major DNA repair pathway in mitochondria is base excision repair (BER) [9], [10]. A great deal has been learned about BER in the context of nuclear DNA damage and repair [11]. Although BER is considered a coordinated process, certain toxic repair intermediates can accumulate under enzyme deficiency and when the amount of DNA lesions overwhelms the repair capacity [11], [12], [13]. One of the repair intermediates is the 5'-deoxyribose phosphate (5'dRp) residue. 5'dRp is mutagenic in cells, resulting in point mutations and deletions, similar to the mutagenic effects of AP sites [14]. 5'dRp is sourced from AP endonuclease (APE1)-catalyzed cleavage 5' to the abasic (AP) site (Fig. 1A). 5'dRp is subject to spontaneous or enzymatic release. DNA polymerases β (pol β) and λ (pol λ) are the primary dRp lyases in BER in the nucleus [15], [16], [17]. In human mitochondria, the major mitochondrial DNA polymerase γ (pol γ) has dRp lyase activities [18]. In addition, pol β has been shown to localize to mitochondria in certain tissues [19], [20], suggesting a potential role of pol β in dRp removal.
Although the cellular level of 5'dRp residues remains unknown due to the lack of specific detection methods, 5'dRp release is considered the rate-limiting step in BER [21], [22], [23], [24]. The biological importance of 5'dRp removal is supported by phenotypes in cell lines and animal models with deficient dRp lyase activities. For instance, the dRp lyase activity of pol β is required to reverse methylating agent hypersensitivity in pol β-null mouse fibroblasts [25], and also contributes to temozolomide resistance in glioma cells [26], [27]. The conditional knock-in mice with a dRp lyase deficient pol β variant (L22P, a gastric cancer-associated variant) exhibit hyperproliferation, DNA double-strand breaks (DSBs), cytosolic DNA-mediated inflammation, and stomach tumors [28], [29], [30]. Moreover, live cell imaging studies have demonstrated that the dRp lyase domain of pol β is crucial for its recruitment to sites of DNA base damage and single-strand breaks after micro-irradiation [31]. Lastly, the critical role of the dRp-lyase activity of the DSB repair factor Ku in non-homologous end joining (NHEJ) corroborates the toxicity of 5'-dRp [32].
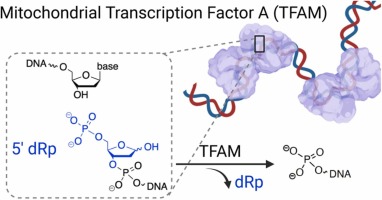
Steady-state kinetic analysis has demonstrated that the catalytic efficiency of pol γ is approximately 20-fold lower than that of pol β in dRp lyase reactions [18]. The relatively slow removal of dRp by pol γ and the tissue-specific mitochondrial localization of pol β motivated us to explore additional proteins with dRp lyase activities in mitochondria. Herein, we examined the ability of removing dRp residues by a major DNA-packaging protein in human mitochondria, mitochondrial transcription factor A (TFAM). TFAM is abundant enough to coat the entire mtDNA and binds specifically to promotor regions to activate mitochondrial transcription [33]. In addition, TFAM has AP lyase activities and participates in processing AP sites in vitro and in cultured cells to facilitate mtDNA turnover [34], [35]. Considering the common Schiff base chemistry-based mechanism of enzymatic AP lyase and dRp lyase reactions (Fig. 2B), we hypothesized that TFAM could act as a noncanonical dRp lyase. Using DNA substrates containing a site-specific dRp residue, we demonstrate that TFAM has dRp lyase activity with a single-turnover rate comparable to that of pol β, albeit slower than pol λ. The lysine residues of TFAM mediate the reaction via Schiff base chemistry. Overall, these results hint at a potential role of TFAM in mtDNA repair.










留言 (0)