A low pH within the stomach is essential for the activation of pepsin, a stomach enzyme which breaks down the protein of ingested food. Acid secretion into the stomach is accomplished by the H+,K+-ATPase (or gastric proton pump), which is located in the plasma membrane of parietal cells of the gastric mucosa. In an electroneutral exchange of K+ ions from the stomach lumen, the H+,K+-ATPase utilises the energy of ATP hydrolysis to pump H+ ions into the stomach (Sachs et al. 1976). Thus, the H+,K+-ATPase plays a crucial role in the digestion process. However, the acidic environment of the stomach provided by the H+,K+-ATPase causes the bacterium Helicobacter pylori, which promotes the formation of peptic ulcers and even stomach cancer, to adopt a coccoid form resistant to antibiotic treatment (Ierardi et al. 2019). The discovery of the mechanism of H+ secretion (Sachs et al. 1976; Forte et al. 1975; Lee and Forte 1978; Wallmark and Mårdh 1979) and the subsequent development of inhibitors (proton pump inhibitors or PPIs), such as omeprazole, can be considered as a revolution in twentieth century medicine, with many millions of patients afflicted with peptic ulcers or gastroesophageal reflux disease being successfully treated (Sachs et al. 2014; Kaunitz 2014).
The gastric H+,K+-ATPase is a member of the P-type ATPase family of enzymes; a large group of integral membrane proteins found in all kingdoms of life which pump small ions, lipids and some other molecules across the membrane in which they are located. They derive the energy they require for the pumping process from the hydrolysis of ATP. They are, therefore, classified as primary active transporters. Transport stoichiometries of the H+,K+-ATPase of 2 H+/2K+ and 1 H+/1K+ per hydrolysed ATP have been reported, but with the stoichiometry accepted to be 1 H+/1K+ at low pH values of the stomach lumen (Reenstra and Forte 1981; Rabon et al. 1982, 1993; Abe et al. 2012; Montes et al. 2011; Yamamoto et al. 2019). The H+,K+-ATPase belongs to the P2C group of the P2 subfamily of P-type ATPases, as does the Na+,K+-ATPase. Both enzymes comprise a catalytic α-subunit and a much smaller β-subunit; in the case of the Na+,K+-ATPase there is a third even smaller subunit with a single membrane-spanning α-helix, which is termed the γ-subunit in kidney cells. The α-subunits of the H+,K+-ATPase and the Na+,K+-ATPase have a high degree of structural homology of approximately 63% (Shin et al. 2009). Therefore, one would expect similarities in the function and regulation of the two enzymes, and hence comparisons in their behaviours to yield useful information.
The primary mode of regulation of acid secretion in the stomach involves recruitment of the H+,K+-ATPase from intracellular tubovesicular elements to the apical membrane surface of stimulated parietal cells upon the ingestion of food (Dunbar and Caplan 2001). Once at the apical membrane surface, the H+,K+-ATPase fuses with the plasma membrane so that it can start pumping H+ ions into the stomach lumen. This poses the question, is there any acute regulation of acid secretion via direct modulation of H+,K+-ATPase transport kinetics? In vitro biochemical studies on the H+,K+-ATPase and comparisons with the Na+,K+-ATPase suggest that this may be the case (Cornelius and Mahmmoud 2003).
Common to both catalytic α-subunits of the H+,K+-ATPase and the Na+,K+-ATPase is a lysine-rich cytoplasmic N-terminal tail. In the case of the Na+,K+-ATPase this tail possesses a tyrosine and a serine residue which are conserved across all vertebrates (Diaz and Clarke 2018). These are potential sites for acute regulation of ion pumping activity via phosphorylation by an Src kinase and a protein kinase C (PKC), respectively (Blayney et al. 2023). Tyr-5 (-10 if the propeptide sequence is included in the numbering) has been identified in cultured kidney cell studies as a Src kinase target (Petrič et al. 2021). Similarly, Ser-11 (-16 if the propeptide sequence is included) has been identified in studies on both purified Na+,K+-ATPase from kidney (Feschenko and Sweadner 1995) and on oocyte homogenates expressing the Na+,K+-ATPase (Beguin et al. 1994) to be a target of PKC. Based on both experimental data and theoretical molecular dynamics simulations, it has been proposed that the regulation occurs via an electrostatic switch mechanism. This involves salt bridge-type interactions between N-terminal positively charged lysine residues and negatively charged lipid headgroups, such as phosphatidylserine, that are weakened by the kinase-induced introduction of the negatively charged phosphate moiety onto the conserved tyrosine and serine residues (Blayney et al. 2023; Lev et al. 2023). Weakening of the salt bridge interactions with the membrane surface is proposed to change the relative stabilities of different conformational states of the Na+,K+-ATPase α-subunit and thus lead to a change in ion pumping activity.
The α1-subunit N-terminus of the H+,K+-ATPase also possesses tyrosine and serine residues in many vertebrate species, but they are not as broadly conserved as in the Na+,K+-ATPase (Diaz and Clarke 2018). Ser-27 (Homo sapiens sequence numbering) appears to be conserved in all placental mammals. Tyr-7 seems to be conserved in all vertebrates except cartilaginous fish, and a tyrosine is also present in position 10 in most mammals (including marsupials), but it is replaced by histidine in some species. Similar to the Na+,K+-ATPase, in vitro experiments (Togawa et al. 1995, 1996; Kanagawa et al. 2000; Fujitani et al. 2003) suggest that some of these residues could be sites of regulatory phosphorylation by kinases in many species. Asano et al. (2000), however, carried out experiments with the H+,K+-ATPase expressed in HEK-293 cells, in which they replaced all of the positively charged lysine residues of the N-terminus with uncharged alanine residues. This had no effect on the stimulation of the enzyme’s steady-state ATPase activity by ATP, K+ or H+. However, these results don’t exclude a role of the N-terminus in determining ion pump kinetics, because there remains the possibility that the N-terminus could play a role in the conformational change of the unphosphorylated enzyme, which is necessary for K+ deocclusion and release into the cytoplasm. That the N-terminus could affect this transition would seem reasonable, considering its location on the cytoplasmic face of the protein. A significant role of the N-terminus of the H+,K+-ATPase α-subunit was in fact reported by Cornelius and Mahmmoud (2003), who found that N-terminal phosphorylation by PKC caused a 40–80% increase in ATPase activity at saturating ATP concentrations, depending on the pH. The number of kinetic studies on partial reactions of the H+,K+-ATPase is much less than that of the Na+,K+-ATPase, but stopped-flow measurements show that in the absence of ATP, the E2 → E1 conformational transition associated with K+ deocclusion to the cytoplasm is 50–100 times faster in the H+,K+-ATPase than in mammalian Na+,K+-ATPase (Rabon et al. 1990; Faraj et al. 2021). This does not exclude, however, a contribution of the E2 → E1 transition to rate determination of the H+,K+-ATPase ion pumping cycle under physiological conditions in the presence of ATP. It is interesting to note here that a higher rate of the E2 → E1 transition is also seen in Na+,K+-ATPase from shark (Squalus acanthias) rectal gland (Khalid et al. 2010). Another similarity between the shark Na+,K+-ATPase and the H+,K+-ATPase is that the α-subunit of both have longer cytoplasmic N-termini with even more lysine residues than that of mammalian Na+,K+-ATPase α1-subunits (Diaz and Clarke 2018). Therefore, it’s possible that the different kinetic behaviours observed with respect to the E2 → E1 conformational transition could be due to differences in the amino acid sequences of the N-termini of the enzymes. No three-dimensional structural data is available on the N-terminus of the H+,K+-ATPase because it was removed prior to crystallisation, probably to facilitate crystal formation due to the disordered nature of the N-terminus (Abe et al. 2018).
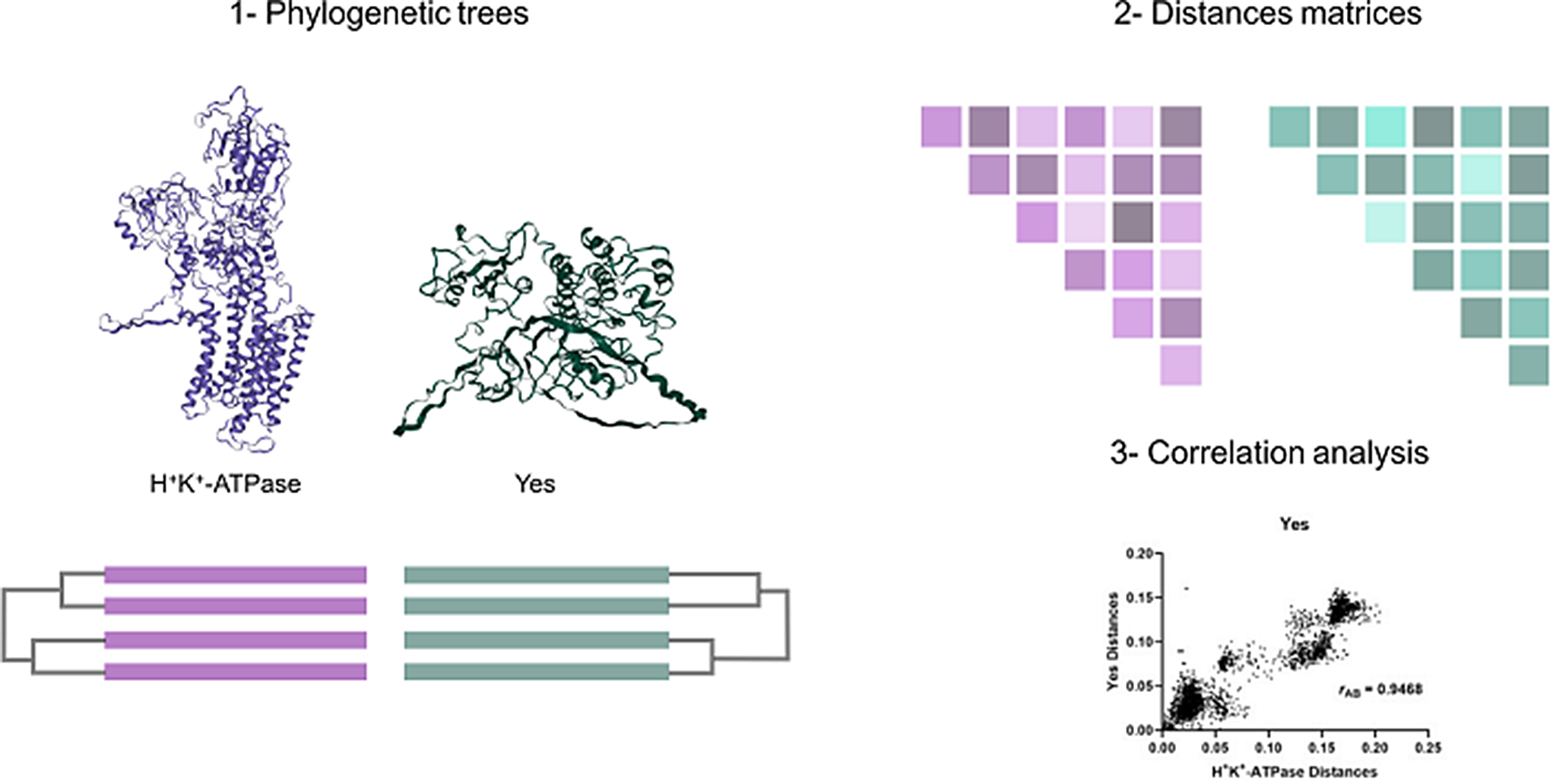
Although experimental data from in vitro experiments support the conclusion that the H+,K+-ATPase could undergo acute regulation via interaction of PKC and the Src kinase with the α-subunit N-terminus, in vivo evidence is lacking. A novel approach to tackling this problem has recently been used on the Na+,K+-ATPase (Blayney et al. 2023). The approach is based on the idea that if two proteins interact with one another, they should be co-evolving, such that if one protein undergoes a mutation, the other protein should undergo a compensating mutation to maintain optimal interaction. Thus, an analysis of the degree of coevolution of the H+,K+-ATPase with protein kinase C and with the Src kinase could potentially provide an indication of whether the enzymes interact with one another in vivo. The bioinformatic method that we use to analyse for co-evolution is called the mirror tree method, because it involves determining the similarity of the two proteins’ phylogenetic trees. Using this method it could be shown that, based on the analysis, the θ and η isoforms of PKC are the isoforms most likely to interact with the Na+,K+-ATPase, and that, of the kinases of the Src family, the Src kinase itself was the most likely reaction partner (Blayney et al. 2023). Thus, not only does the method allow one to analyse whether kinase regulation likely occurs in vivo, if it does, the method also allows the most likely kinase isoform involved to be identified. This is then very useful information that could be used to guide future experimental investigations.
Here we present mirror tree analyses of the coevolution of the α1-subunit of the H+,K+-ATPase with all 10 isoforms of PKC (α, β1, β2, γ, δ, ε, η, θ, ζ and λ(ι)) and all 9 members of the Src kinase family (Src, Yes, Fyn, Fgr, Lck, Hck, Blk, Lyn and Frk). We also present an expanded data set of amino acid sequence alignments of the H+,K+-ATPase α1-subunit to determine the degree of conservation of possible regulatory kinase targets. The results support the conclusion that the H+,K+-ATPase could undergo acute regulation by kinase phosphorylation in many animal species.

留言 (0)