
記住我
Colorectal cancer (CRC) is the third most commonly diagnosed cancer and the second cause of cancer-related deaths worldwide (Sung et al., 2021). The highest incidence of CRC can be found in Europe, North America and Oceania, which is likely due to lifestyle factors, i.e., less physical activity, consumption of high-calorie-dense food, and smoking (Center et al., 2009). Indeed, the latter have been identified as CRC risk factors, next to age, gender, family history, colitis, alcohol consumption, high consumption of red and processed meat, obesity and diabetes (Taylor et al., 2010; Chan et al., 2011; Fedirko et al., 2011; Jiang et al., 2011; Jess et al., 2012; Ma et al., 2013). On the other hand, there are also elements that are associated with a lower chance to develop CRC, such as physical activity, a healthy diet, removal of precancerous lesions, hormone replacement therapy, and aspirin (Brenner et al., 2011; Bosetti et al., 2012; Boyle et al., 2012; Lin et al., 2012; Veettil et al., 2021). However, the reported lower incidence of CRC in Asia and Africa might also be attributed to poor access to healthcare and screening tools (Pourhoseingholi, 2012; Awedew et al., 2022).
Treatment of CRC is based on tumor- (e.g., tumor size/progression, presence and localization of metastases) and patient-related factors (e.g., prognosis, general health, age) (Mármol et al., 2017). Generally, the standard course of treatment for CRC is surgery (Kuipers et al., 2015). Neoadjuvant (before surgery) and adjuvant (after surgery) therapies mostly involve chemotherapy, radiotherapy or a combination of both (Brenner et al., 2014; Kuipers et al., 2015). However, because treatment is adapted to the patient during the course of treatment, it is difficult to define the percentage of patients receiving a certain treatment course.
Radiotherapy is an important treatment option for CRC, but is also associated with acute and/or chronic toxicity (Häfner and Debus, 2016; Segers et al., 2019). Acute symptoms of pelvic radiotoxicity are diarrhea, nausea, fatigue and abdominal pain (Hauer-Jensen et al., 2014). Chronic pelvic radiotoxicity is associated with changes in intestinal transit, malabsorption, impaired gut motility, fistula formation, intestinal obstruction and perforation (Hauer-Jensen et al., 2014; Segers et al., 2019). These side effects can be so severe that treatments need to be stopped temporarily or even permanently. The risk to develop radiation-related side effects is partly therapy- (e.g., radiation dose, fraction, site, and concomitant treatments) and partly patient-related (e.g., sex, age, genetic susceptibility, and smoking) (Bentzen and Overgaard, 1994; Andreassen and Alsner, 2009). In addition, radiotherapy can disrupt the gut microbiome, which in turn can influence radiotherapy efficacy by hindering the ability to repair radiation-induced intestinal damage (Liu et al., 2021).
Other emerging therapeutics are also being applied on CRC. As cancer cells are able to evade the immune system, there is a scientific ground to target the tumor by blocking these immune evasion mechanisms or activating the immune system (Kim and Cho, 2022). Furthermore, the presence of T-cells in the CRC tumor microenvironment is associated with better prognosis, indicating that targeting T-cells might be useful for this cancer type (Galon et al., 2006; Galon et al., 2007). Therefore, the use of immunotherapy has been explored for the treatment of CRC (Ganesh et al., 2019). More specifically, immune checkpoint blockers (ICBs) that enhance T-cell activation by blocking the co-inhibitory T-cell receptor programmed cell death 1 (PD-1) or its ligand (PD-L1) or cytotoxic T lymphocyte antigen 4 (CTLA-4) have shown promise. Based on the KEYNOTE-177 and CheckMate 142 clinical trials, the anti-PD-1 molecules pembrolizumab and nivolumab, and the combination of nivolumab and the anti-CTLA-4 molecule ipilimumab have been approved by the Food and Drug Administration (FDA) for CRC treatment (Overman et al., 2017; Diaz et al., 2022; Lenz et al., 2022). An important predictive marker for ICB efficacy is microsatellite instability (MSI). Microsatellites are short tandem repeat DNA sequences that are more susceptible to replication defects and are normally corrected by mismatch repair (MMR) (De’ Angelis et al., 2018). An incorrect repair by the MMR system results in the MSI phenotype with the generation of frameshift mutations and synthesis of neoantigens, making cancer cells more easily detectable by the immune system (Maby et al., 2016). Based on their frequency, three different microsatellite statuses have been characterized: high MSI (MSI-H), low MSI (MSI-L) and microsatellite stability (MSS) (Bonneville et al., 2020). Patients who are MSI-H can also be identified as MMR deficient (dMMR), whereas patients who are MSI-L or MSS can be identified as MMR proficient (pMMR) (Sinicrope and Yang, 2011). ICBs are only applicable for MSI-H CRC patients, since MSI-L and MSS CRC patients are unresponsive to ICBs due to the lack of tumor mutational burden, little tumor-infiltrating lymphocytes (TILs), low PD-L1 expression on tumor cells and low IFN-γ expression (Galon et al., 2006; Le et al., 2017; Ganesh et al., 2019). However, only 15% of all CRC patients are MSI-H, suggesting that only a small subset of patients could benefit from immunotherapy (Boland et al., 1998). Furthermore, increasing evidence suggests that the gut microbiome is another important factor influencing anti-tumor responses and ICB efficacy in CRC (Hou et al., 2022).
Besides the efficacy-related problems, ICBs induce the activation of T-cells and reduce the functions of regulatory T-cells (Tregs), which can lead to overstimulation of the immune system (Ramos-Casals et al., 2020). This can result in immunotherapy-related adverse events in all organs, including the gastrointestinal (GI) (e.g., colitis, hepatitis), dermatologic (e.g., alopecia, psoriasis, vitiligo), cardiovascular (e.g., myo- or endocarditis, cardiomyopathy), pulmonary (e.g., alveolitis, pneumonitis), neurologic (e.g., encephalitis, meningitis), systemic (e.g., cytokine release syndrome) and endocrine (e.g., thyroiditis, adrenitis) systems (Martins et al., 2019; Ramos-Casals et al., 2020; Chhabra and Kennedy, 2021).
This brief introduction highlights the importance of radiotherapy and immunotherapy as treatment options for CRC. However, it also shows that both treatments have drawbacks concerning inconsistency and side effects, with increasing evidence pointing towards the gut microbiome as a driver for these issues. In this review, we will focus on the gut microbiome and summarize how it influences radio- and immunotherapy efficacy and related side effects in CRC. Noteworthy, the efficacy and toxicity of chemotherapy, one of the most common treatment options for CRC (McQuade et al., 2017; Kumar et al., 2023), are also affected by the gut microbiome. We consider this out of scope and refer to extensive reviews (Kalasabail et al., 2021; Yu et al., 2022; Liu et al., 2023).
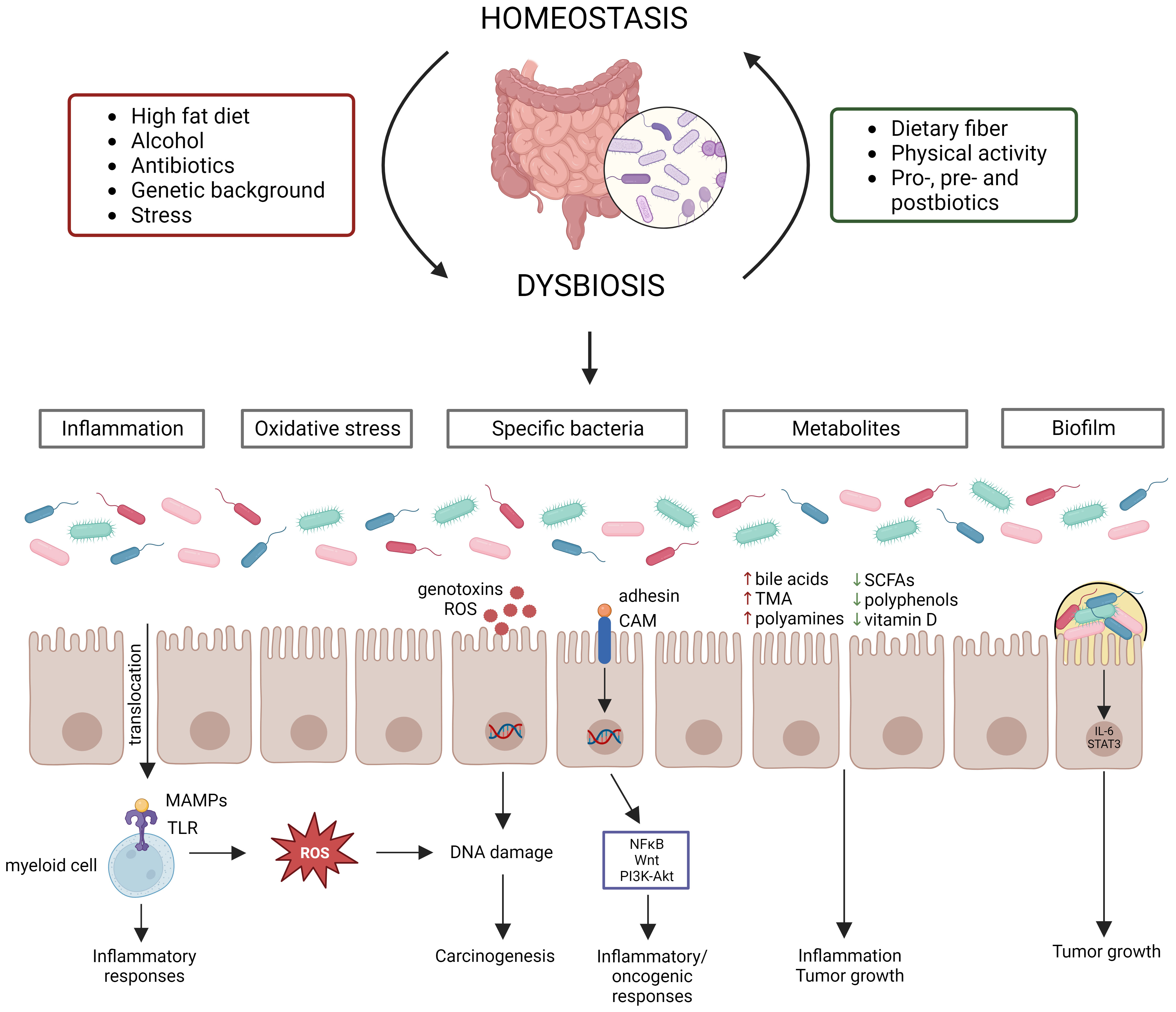
2 The gut microbiome in CRCA typical healthy gut microbiome contains trillions of microbes (Cresci and Bawden, 2015), covering between 300-1000 bacterial species, with Bacteroidetes and Firmicutes being the most abundant phyla (Lozupone et al., 2012). Actinobacteria, Proteobacteria and Verrucomicrobia are also present in relatively high abundances (Lozupone et al., 2012). The gut microbiome has three main functions: structural (maintenance of structural integrity of gut mucosal barrier), protective (co-relationship with the immune system to fight against invading microorganisms) and metabolic (e.g., participation in digestive processes, production of metabolites or other molecules) (Jandhyala et al., 2015; Rebersek, 2021). The host and its microbiome are one unit that co-evolve with each other and its composition can be influenced by multiple factors (e.g., diet, genetic background, stress, physical activity, and anti-, pro-, pre-, and postbiotic intake) (Figure 1) (Deschasaux et al., 2018; Leeming et al., 2019; Clooney et al., 2021). Disruption of the intestinal bacterial homeostasis is defined as dysbiosis, which is characterized by an altered diversity and abundance of the associated microbiota that can negatively impact the immune system (DeGruttola et al., 2016; Toor et al., 2019; Berg et al., 2020). Dysbiosis has also been linked to multiple diseases, such as neurological diseases, inflammatory diseases (e.g., inflammatory bowel disease (IBD)) and cancer (Hullar et al., 2014; Nishida et al., 2018; Sun and Shen, 2018).
Figure 1 Gut microbiome homeostasis can be disrupted by various factors leading to dysbiosis, which contributes to CRC development via distinct mechanisms. Multiple factors influence the gut microbiome composition, establishing homeostasis or dysbiosis. Dysbiosis disrupts the intestinal barrier, resulting in translocation of bacteria and their products, which starts an inflammatory cascade. Moreover, certain bacteria produce reactive oxygen species (ROS) or genotoxins, while other bacteria activate inflammatory/oncogenic pathways. Chronic inflammation also results in the production of ROS, leading to DNA damage. Dysbiosis can also induce changes in metabolite levels, which contribute to inflammation and tumor growth. Lastly, biofilms can be formed due to dysbiosis, which also contribute to tumor growth.
Evidence from several studies has shown that dysbiosis can contribute to CRC carcinogenesis through multiple mechanisms, such as (1) pathogenic bacteria and their toxic products, (2) inflammation, (3) oxidative stress, (4) metabolites, and (5) biofilm formation (Figure 1). Dysbiosis is associated with a loss of protective bacteria, such as Bifidobacterium animalis, Clostridium butyricum and Streptococcus thermophiles, and an enrichment of cancer-promoting bacteria, such as Fusobacterium nucleatum, Bacteroides fragilis, Escherichia coli, Streptococcus bovis, Enterococcus faecalis and Peptostreptococcus anaerobius (Feng et al., 2015; de Almeida et al., 2018; Wassenaar, 2018; Long et al., 2019; Sun et al., 2019; Cheng et al., 2020; Deng et al., 2020; Li et al., 2021; Liu et al., 2021). It is to be noted that the contribution to CRC cannot be attributed to a sole bacterium, but multiple bacteria elicit negative effects that surpass those of the beneficial bacteria (Cheng et al., 2020). Several mechanisms by which these cancer-promoting bacteria can contribute to CRC development and progression have been proposed. For instance, F. nucleatum contains the surface virulence factors FadA, which activates inflammatory/oncogenic responses via NFκB and Wnt signaling, and Fap2, which protects CRC cells from immune attack (Sun et al., 2019). The B. fragilis toxin has also been shown to activate NFκB and Wnt signaling thereby promoting tumor cell proliferation and inducing metastasis (Cheng et al., 2020). The E. coli B2 phylotype can produce the genotoxin colibactin, which induces dsDNA breaks in the intestinal epithelial cells (Wassenaar, 2018). S. bovis has been shown to recruit CD11b+TLR4+ cells, which could promote a pro-tumor microenvironment (Deng et al., 2020). E. faecalis contributes to CRC carcinogenesis by producing reactive oxygen species (ROS), which induce DNA damage, and metalloproteases, which compromise barrier integrity and contribute to inflammation (de Almeida et al., 2018). P. anaerobius contains the surface protein PCWBR2, which preferentially interacts with CRC cells via α2/β1 integrin, after which the PI3K-Akt pathway is activated, eventually generating a pro-inflammatory response (Long et al., 2019). The detrimental effect of the CRC gut microbiome has been demonstrated by transplanting fecal samples from CRC patients to germ-free and conventional mice, leading to increased tumor proliferation, alteration of the gut microbiome, increased expression of pro-inflammatory genes and increased immune cell infiltration (Wong et al., 2017). The changed microbiome composition can affect the intestinal barrier, which results in translocation of microbiota and their products to extraintestinal sites, such as mesenteric lymph nodes, kidney, liver, spleen and bloodstream (Berg, 1999; Maisonneuve et al., 2018; Zou et al., 2018). Furthermore, bacterial translocation has been shown to contribute to CRC metastasis (Keramidaris et al., 2013). The translocated bacteria are recognized by immune cells via Toll-like receptors (TLRs), which induces the expression of cytokines, starting a pro-inflammatory cascade (Sánchez-Alcoholado et al., 2020). Inflammation induces DNA damage in intestinal cells, dysregulates anti-tumor immune responses and alters the gut microbiome composition, thereby contributing to CRC development (Nagao-Kitamoto et al., 2022). The idea that inflammation contributes to CRC carcinogenesis is supported by the observation that IBD patients have an increased risk of 10 – 15% to develop CRC (Loddo and Romano, 2015). Furthermore, dysbiosis is associated with altered levels of gut microbial metabolites, such as short-chain fatty acids (SCFAs), bile acids, trimethylamine (TMA), polyamines, polyphenols and vitamins (Bhat and Kapila, 2017; Zou et al., 2018). SCFAs, such as butyrate and propionate, can block inflammation by promoting Treg differentiation and stimulating macrophages and T-cells to produce IL-10 and TGF-β (Zou et al., 2018). The secondary bile acids deoxycholic acid and lithocholic acid promote CRC development by activating β-catenin and epidermal growth factor signaling and promoting cancer invasion and MAPK signaling, respectively (Baek et al., 2010; Ha and Park, 2010). TMA is further processed to trimethylamine-N-oxide, which is associated with CRC development, although its mechanism is still unclear (Xu et al., 2015). Polyamines are upregulated in CRC cells and stimulate tumor growth and immune evasion (Casero and Marton, 2007; Hayes et al., 2014; Novita Sari et al., 2021). Polyphenols show a protective effect by affecting inflammation, gut microbiota, epigenetics and mRNA expression (Ding et al., 2020). Vitamin D also elicits anticancer effects by influencing inflammation, apoptosis and Wnt/β-catenin signaling (Javed et al., 2020). The chronic inflammation induced by dysbiosis can result in the release of ROS, which can induce DNA damage of intestinal epithelial cells, eventually contributing to CRC development (Sorolla et al., 2021). Furthermore, it is hypothesized that certain bacteria can directly cause DNA damage (Artemev et al., 2022). For instance, E. coli has been shown to contribute to CRC development by downregulating DNA MMR (Sobhani et al., 2013). Dysbiosis can also contribute to the formation of biofilms, which have been linked to CRC development (Chew et al., 2020). Biofilms reduce the epithelial E-cadherin, resulting in disruption of the intestinal barrier, and stimulate IL-6/STAT3 signaling, inducing intestinal epithelial cell proliferation and tumor growth (Dejea et al., 2014; Johnson et al., 2015). In addition, biofilms can contribute to the formation of polyamine metabolites (Johnson et al., 2015).
2.1 The gut microbiome as a biomarker in CRCSince the gut microbiome is altered in CRC patients, the gut microbiome is proposed as a screening, prognostic or predictive biomarker (Rebersek, 2021). For instance, it has already been shown that the gut microbiome differs between dMMR and pMMR cancer patients (Tahara et al., 2014; Mima et al., 2016; Gopalakrishnan et al., 2018; Hale et al., 2018; Matson et al., 2018; Routy et al., 2018; Xu et al., 2020; Kang et al., 2021; Jin et al., 2022). In one study, it was discovered that dMMR CRC patients showed a higher alpha diversity than pMMR CRC patients (Jin et al., 2022). Furthermore, dMMR CRC patients carried a higher abundance of the Fusobacteria, Firmicutes, Verrucomicrobia and Actinobacteria phyla and Fusobacterium, Akkermansia, Bifidobacterium, Faecalibacterium, Streptococcus and Prevotella genera. In contrast, pMMR CRC patients showed a higher abundance of Proteobacteria and more specifically the Serratia, Cupriavidus and Sphingobium genera. Based on the microbiome composition, Jin et al. predicted that the dMMR status could be associated with biosynthetic/metabolic pathways of glycan, vitamins and nucleotides, cell growth and death pathways, and genetic replication and repair pathways. The pMMR status was predicted to be associated with lipid, terpenoid, polyketone and amino acid metabolic pathways and membrane transport pathways. Other studies with CRC patients corroborated these observations and showed enrichment in Fusobacteria and Bacteroidetes, and reduction in Firmicutes and Proteobacteria in dMMR patients (Tahara et al., 2014; Mima et al., 2016; Hale et al., 2018; Xu et al., 2020; Kang et al., 2021). The pMMR patients also showed altered metabolic pathways of glycerol and phospholipid (Xu et al., 2020).
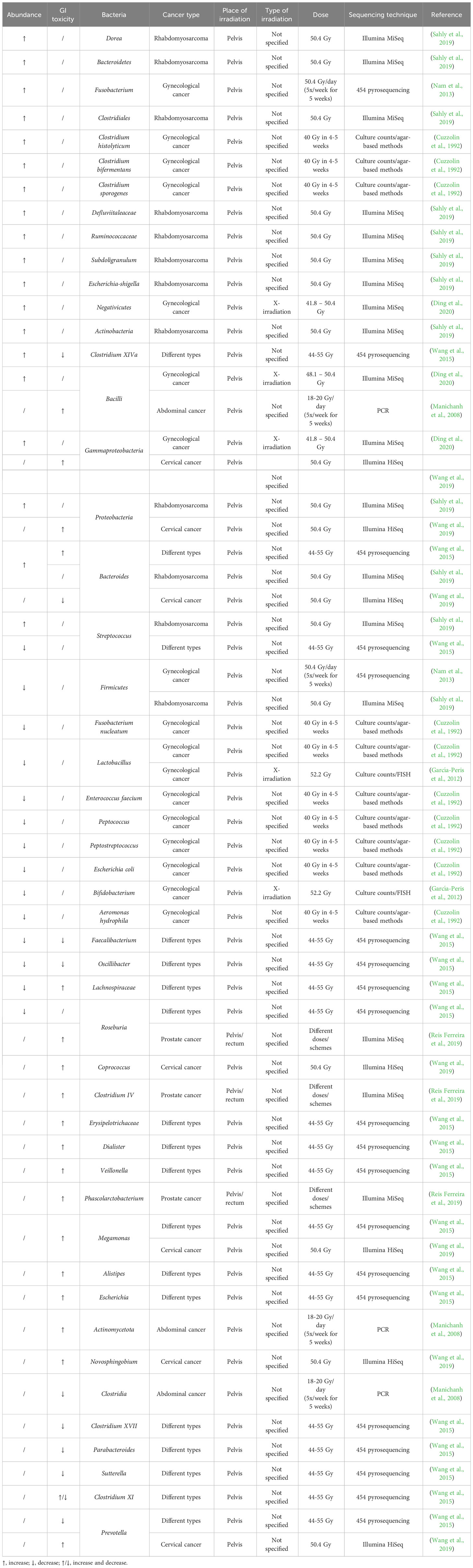
3 Radiotherapy-induced changes in the gut microbiome and its influence on toxicityDespite its importance, the effect of irradiation on the human gut microbiome has been poorly studied (Table 1). An early study investigating the effect of pelvic radiotherapy on the gut microbiome in gynecological cancer patients discovered a decreased abundance of E. coli, Aeromonas hydrophila, Peptococcus spp., Peptostreptococcus spp., F. nucleatum, Enterococcus faecium and Lactobacillus, and an increased abundance of Clostridium histolyticum, Clostridium bifermentans and Clostridium sporogenes after radiotherapy (Cuzzolin et al., 1992). Other studies on gynecological cancer patients found that pelvic radiotherapy also decreased the abundance of the Firmicutes phylum and the Lactobacillus and Bifidobacterium genera, and increased the abundance of the Fusobacterium phylum and Gammaproteobacteria, Bacilli and Negativicutes classes (García-Peris et al., 2012; Nam et al., 2013; Ding et al., 2020). Another study investigated the effect of pelvic irradiation on the gut microbiome of 11 patients with different cancer types and discovered that irradiation induced an increase in Bacteroides and Clostridium XIVa and a decrease in Lachnospiraceae, Faecalibacterium, Roseburia, Oscillibacter and Streptococcus (Wang et al., 2015). When pediatric patients with rhabdomyosarcoma near the pelvic area received pelvic radiotherapy, the abundance of Firmicutes decreased whereas the abundance of Proteobacteria, Actinobacteria and Bacteroidetes increased (Sahly et al., 2019). Furthermore, the abundance of Defluviitaleaceae, Ruminococcaceae, Clostridiales, Bacteroides, Streptococcus, Dorea, Bacteroides, Subdoligranulum, Escherichia-Shigella increased after radiotherapy. When investigating the role of the gut microbiome on radiation-induced diarrhea in 10 patients with abdominal cancer, it was discovered that Actinobacteria and Bacilli were associated with radiation-induced diarrhea, whereas Clostridia was not (Manichanh et al., 2008). Before pelvic irradiation of 11 patients with different cancer types, patients who developed post-radiation diarrhea had increased abundances of Bacteroides, Dialister, Veillonella and decreased abundances of Clostridium XI, Clostridium XVII, Faecalibacterium, Oscillibacter, Parabacteroides and Prevotella in their fecal samples (Wang et al., 2015). Post-irradiation, the relative abundances of Clostridium XI together with Alistipes, Bacteroides, Erysipelotrichaceae, Escherichia, Lachnospiracea and Megamonas, were significantly higher in patients who developed diarrhea, whereas Clostridium XIVa and Sutterella were significantly lower. Reis Ferreira et al. investigated the effect of the gut microbiome on radiation enteropathy in prostate cancer patients and discovered that Clostridium IV, Roseburia and Phascolarctobacterium were associated with radiation enteropathy (Reis Ferreira et al., 2019). They also showed that homeostatic intestinal mucosa cytokines related to microbiota regulation and intestinal barrier maintenance were reduced in patients with radiation enteropathy. Cervical cancer patients with radiation enteritis (RE) showed a higher abundance of Proteobacteria, Gammaproteobacteria, Megamonas, Novosphingobium and Prevotella and a decreased abundance of Bacteroides compared to patients without RE (Wang et al., 2019). Furthermore, patients who would later develop RE showed a significantly higher abundance of Coprococcus, indicating that this could be a possible biomarker predicting the chance of RE.
Table 1 Radiation-induced up- or downregulation of bacteria and their correlation with radiotoxicity in cancer patients.
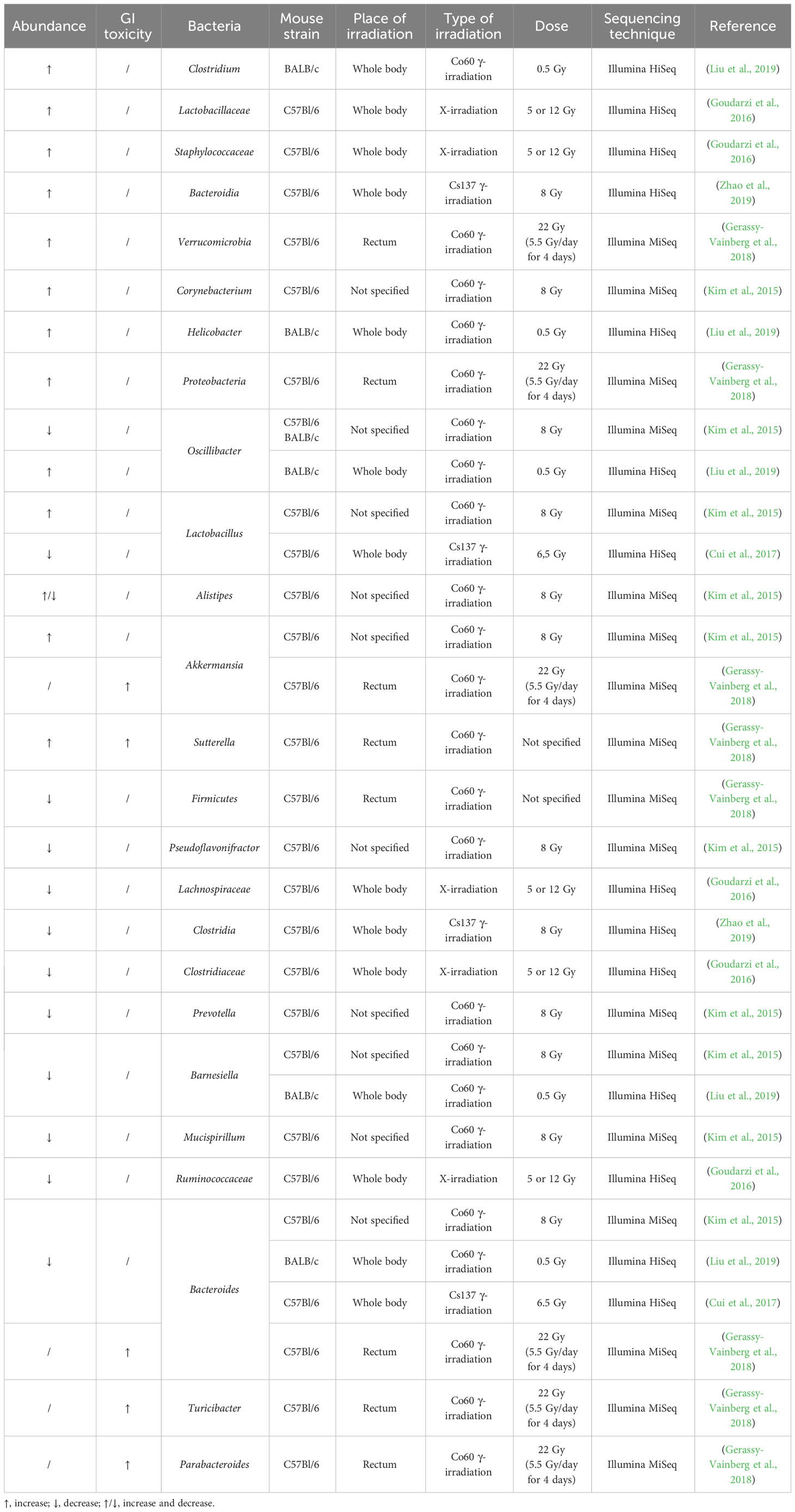
The effect of irradiation on the gut microbiome has been more extensively investigated in mice, although only in healthy mice (Table 2). Acute pelvic irradiation of mice induced intestinal damage and inflammation, which resulted in loss of tight junctions, eventually leading to translocation of bacteria to mesenteric lymph nodes and dysbiosis (Segers et al., 2021). Radiation-induced dysbiosis can aggravate intestinal inflammation, which was shown by an increased expression of IL-1β (Gerassy-Vainberg et al., 2018). Members of the Ruminococcaceae, Lachnospiraceae and Porhyromonadaceae families could be identified as markers of dysbiosis (Segers et al., 2021). Gamma irradiation of mice led to increased proportions of Alistipes, Lactobacillus and Akkermansia, but reduced proportions of Barnesiella, Prevotella, Bacteroides, Oscillibacter, Pseudoflavonifractor and Mucispirillum in the large intestine (Kim et al., 2015). In the small intestine, irradiation led to an increase in Corynebacterium and decrease in Alistipes. In addition, Gerassy-Vainberg et al. exposed mice to rectal irradiation, leading to an increased abundance of Proteobacteria and Verrucomicrobia and a decreased abundance of Firmicutes (Gerassy-Vainberg et al., 2018). When exposed to low-dose irradiation, mice showed an increased abundance of Clostridium, Helicobacter and Oscillibacter and a decreased abundance of Bacteroides and Barnesiella (Liu et al., 2019). The latter study also showed that irradiated mice had perturbed metabolite levels (e.g., downregulation of glucose, pyruvic acid, pinitol, and upregulation of hydroquinone, octadecanol and O-phosphoserine), which were predicted to be involved in glucagon signaling, central carbon metabolism and type II diabetes. Goudarzi et al. investigated the effect of X-ray radiation on mice and discovered that irradiation led to an increased abundance of Lactobacillaceae and Staphylococcaceae, and a decreased abundance of Lachnospiraceae, Ruminococcaceae and Clostridiaceae (Goudarzi et al., 2016). The metabolomic data revealed statistically significant changes in the microbiota-derived products, such as pipecolic acid, glutaconic acid, urobilinogen and homogentisic acid. In addition, significant changes were detected in bile acids (e.g., taurocholic acid and 12-ketodeoxycholic acid), which may be associated with an altered abundance of Ruminococcus gnavus that is able to transform bile acids (Devlin and Fischbach, 2015; Goudarzi et al., 2016). An increased abundance of Bacteroidia and a decreased abundance of Clostridia could still be observed in irradiated mice ten months after exposure, advocating long-term effects of irradiation (Zhao et al., 2019). The gut microbiome can also influence the risk of side effects, corroborated by the higher survival rates of mice that received antibiotics before radiotherapy (Cui et al., 2017). Members of Akkermansia, Bacteroides, Parabacteroides, Sutterella and Turicibacter were more abundant in mice who developed radiation proctitis (Gerassy-Vainberg et al., 2018). Furthermore, irradiation of germ-free (GF) mice that were inoculated with fecal material from previously irradiated mice led to worse irradiation-induced damage compared to irradiation of GF mice that were inoculated with fecal material from naïve mice (Gerassy-Vainberg et al., 2018).
Table 2 Radiation-induced up- or downregulation of bacteria and their correlation with radiotoxicity in healthy mice.
It is proposed that the gut microbiome can influence radiotherapy response, for which some markers have already been defined (Liu et al., 2021). Autophagy has been shown to be related to radiosensitivity/radioresistance of the tumor. Inhibition of autophagy induces radioresistance whereas induction of autophagy induces radiosensitivity (Kuwahara et al., 2011). Fusobacterium nucleatum has been shown to activate autophagy, leading to chemoresistance (Yu et al., 2017). Up until now, no studies have shown the effect of the gut microbiome on radioresistance via autophagy. Furthermore, it has been shown that other factors such as the time when radiation is given can affect the gut microbiome leading to differences in radiosensitivity (Cui et al., 2016; Chan et al., 2017). Fasting-induced adipose factor (FIAF) is a microbiota-regulated protein that has been related to the radiosensitivity of endothelial cells and lymphocytes, and could be used as a protector for radiotoxicity (Crawford and Gordon, 2005). Bacteroides thetaiotaomicron and Enterococcus faecalis increase the FIAF production, whereas Escherichia coli decreases the FIAF production (Grootaert et al., 2011). However, there is currently no information available about the influence of the gut microbiome on radiotherapy efficacy as a stand-alone treatment.
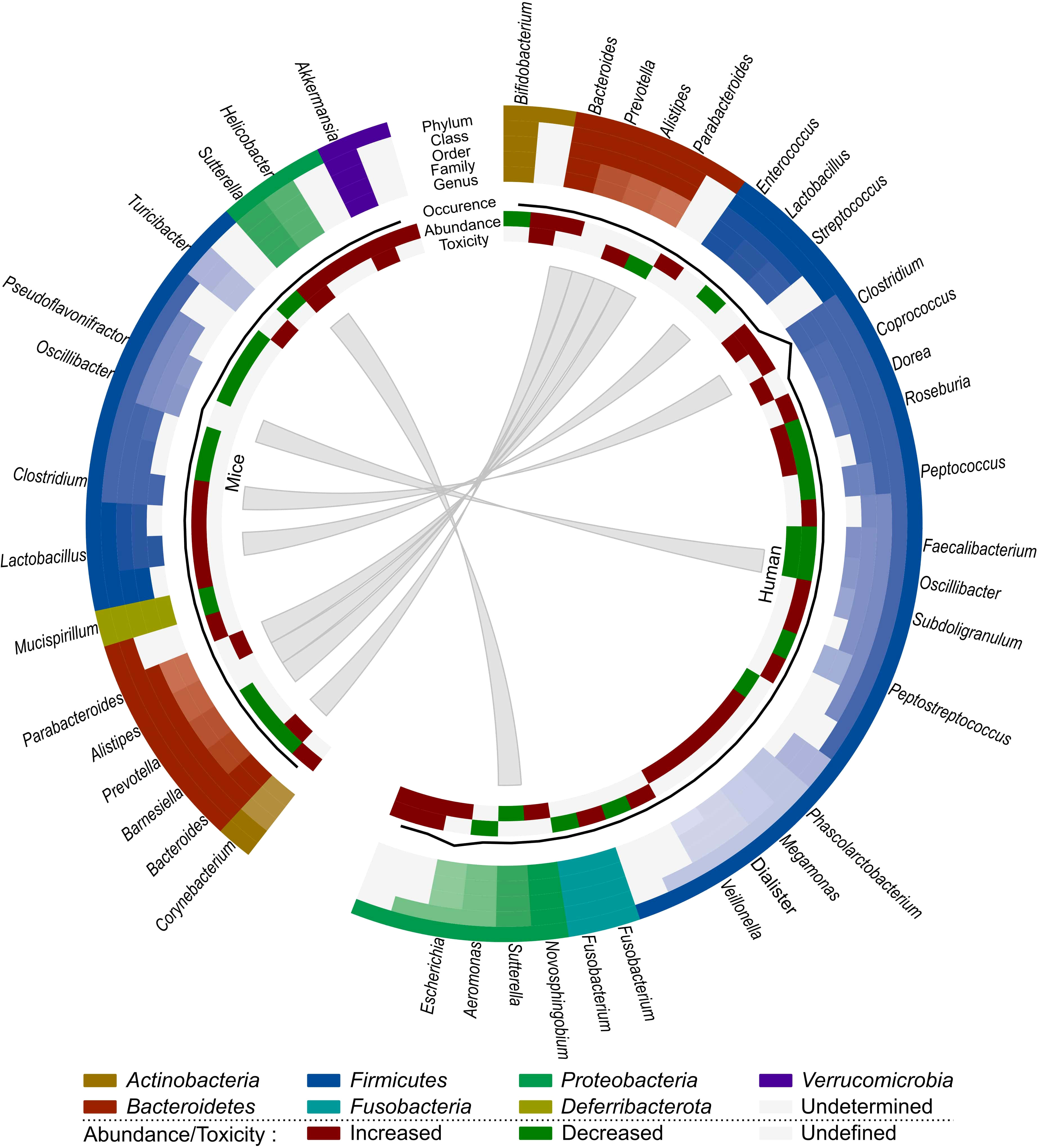
We have opted to list the outcome of different studies exhaustively in order to highlight the complex effects of irradiation on the gut microbiome. Based on all these observations, we can conclude that the presence of Bacilli, Negativicutes, Lachnospiraceae, Coprococcus, Escherichia and Alistipes seem to be related to radiation-induced toxicity in human cancer patients, whereas Akkermansia, Bacteroides, Sutterella, Parabacteroides and Turicibacter are associated with radiation-induced toxicity in mice (Figure 2). However, Sutterella and Parabacteroides seem to be associated with less toxicity in cancer patients. In addition, Faecalibacterium and Oscillibacter reduce toxicity and the contribution of Bacteroides remains unclear. Nevertheless, comparison of mice and human data is difficult, since besides intrinsic differences in their microbiome, there is a lack of data about the role of the microbiome in radiotherapy-induced toxicity in cancer-bearing mice. Moreover, the impact of the gut microbiome on radiotherapy efficacy remains unknown in both human and mice, presenting an intriguing area for further investigation.
Figure 2 Comparison of the effect of radiotherapy on the gut microbiome of mice and humans. Each distinct phylum is represented by a separate color, as are distinct taxonomic classes, orders, families and genera. Similar observations are connected. Additional information is provided for each observation, including its occurrence in literature (line, scale maximum is 7), (relative) abundance of member and toxicity (increased: red; decreased: green; undefined: grey).
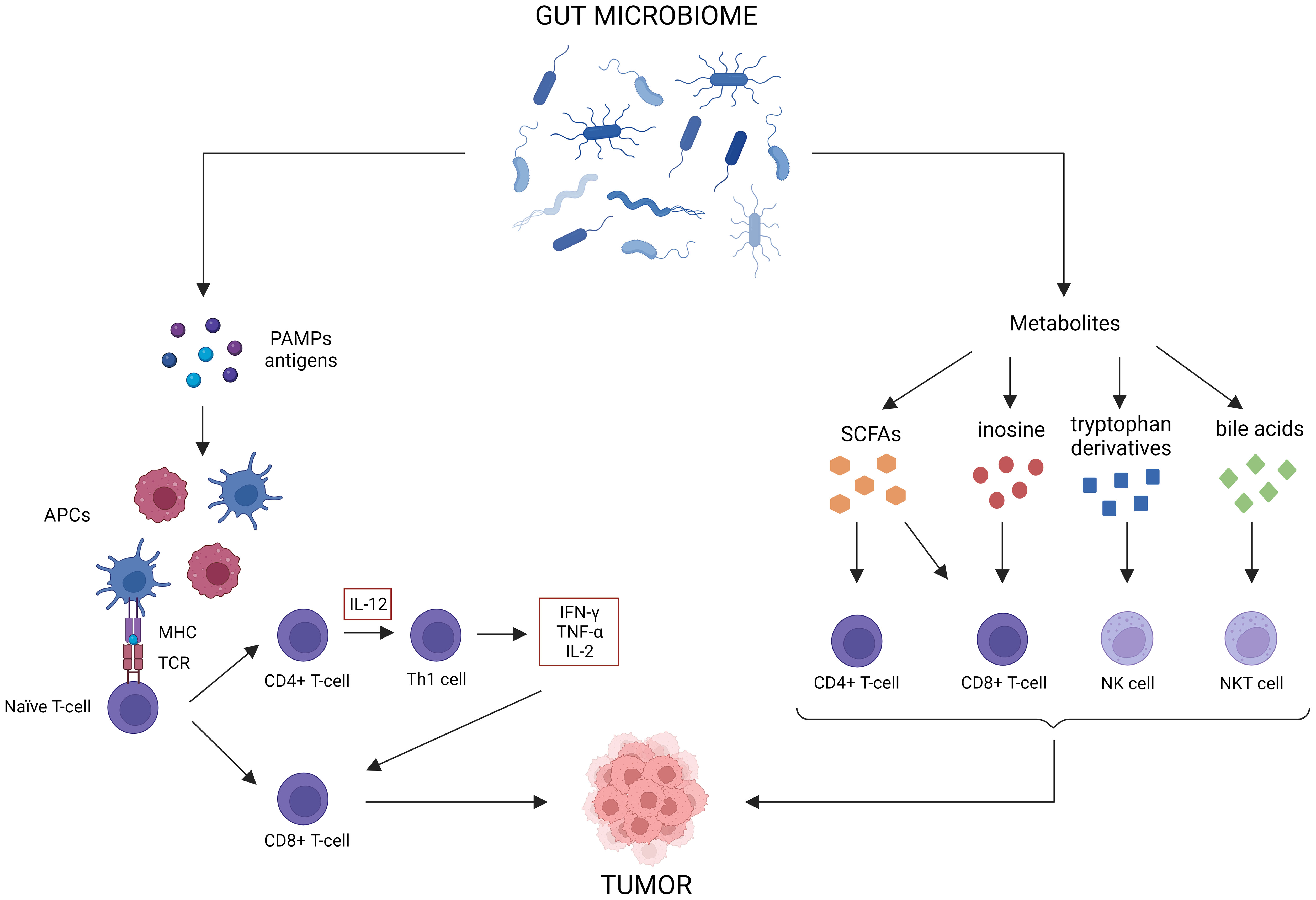
4 The influence of the gut microbiome on anti-tumor responses and immunotherapy efficacyFor CRC, ICBs such as anti-PD-L1, anti-PD-1 or anti-CTLA-4, are only indicated for the treatment of dMMR cancer (Ganesh et al., 2019). However, only 15% of all CRC patients are dMMR, meaning only a small subset of patients will benefit from immunotherapy as a monotherapy (Boland et al., 1998). Most patients will need additional treatment to achieve an optimal response. As the gut microbiome is involved in maturation of both the innate and the adaptive immune system, perturbation can result in aberrant immune responses that can contribute to multiple GI disorders, such as IBD and cancer (Zheng et al., 2020; Jain et al., 2021). Likewise, the abundance of some bacteria (e.g., Bacteroidetes, Akkermansia, Lactobacillus) are positively correlated to antitumor immunity, whereas others are negatively correlated (e.g., Firmicutes, Proteobacteria, Parabacteroides) (Xia et al., 2020). The gut microbiome can elicit antitumor immune responses via different mechanisms (Figure 3) (Park et al., 2020). For instance, some microorganisms contain pathogen-associated molecular patterns that can activate antigen-presenting cells (APCs) via pattern recognition receptors (Panda et al., 2018). This induces the activation of CD4+ and CD8+ T-cells, which influences cytokine expression (TNF-α, IFN-γ, IL-2) and stimulates tumor cell killing (Li et al., 2019). Microbial and tumor cell products may also share antigen sequences, leading to T-cell cross-reactivity between these antigens, which induces antigen-specific immune responses known as molecular mimicry (Baruch et al., 2021). Furthermore, multiple gut microbiota metabolites can elicit antitumor effects. For instance, SCFAs have been shown to reduce colon liver metastases in mice models, activate cytotoxic CD8+ T-cells and their memory potential and stimulate and differentiate CD4+ T-cells (Park et al., 2016; Bachem et al., 2019; Ma et al., 2020; He et al., 2021). Inosine has been shown to act as a carbon source for CD8+ T-cells and stimulates T-cell proliferation and differentiation while enhancing sensitivity to ICBs (Wang et al., 2020). Tryptophan derivatives can stimulate the cytolytic activity of natural killer (NK) cells (Shin et al., 2013). Finally, the gut microbiome can also modify bile acids, which are able to activate antitumor immune cells, such as natural killer T-cells (NKT) (Sipe et al., 2020).
Figure 3 Antitumor immune effects of the gut microbiome. The gut microbiome possesses multiple immune-related antitumor mechanisms. First of all, certain microorganisms release pattern-associated molecular patterns (PAMPs), which activate antigen-presenting cells (APCs) to stimulate CD4+ and CD8+ T-cell responses. Molecular mimicry between tumor and microbial antigens can contribute to these responses as well. Lastly, gut microbiome metabolites, such as short-chain fatty acids (SCFAs), inosine, tryptophan derivatives and bile acids, can activate antitumor immune cells.
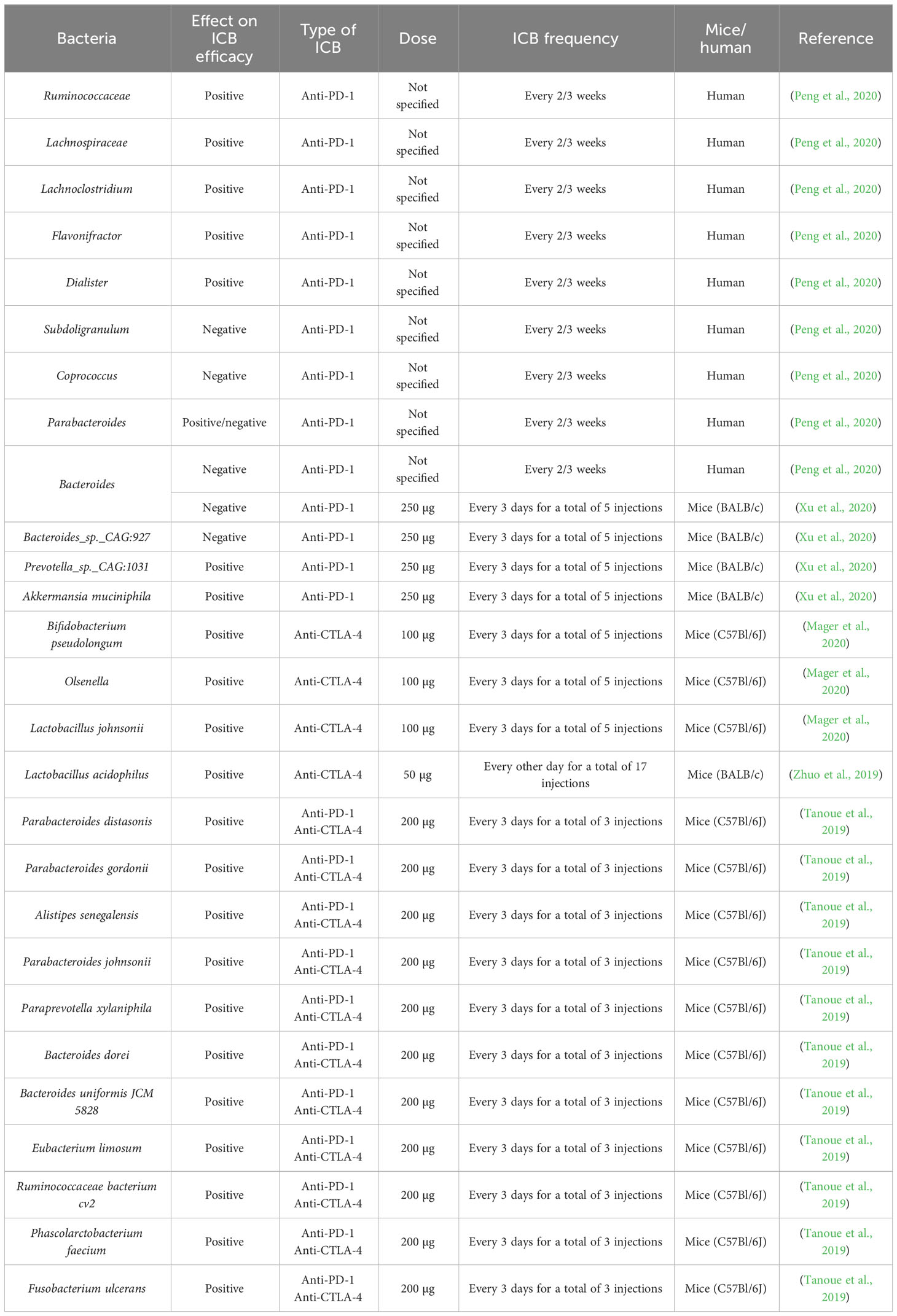
Although there is a lot of knowledge on the influence of the gut microbiome on immunotherapy efficacy for several cancer types, there is limited information for CRC, which is surprising considering its involvement with the microbiome (Table 3). A study involving a cohort of GI cancer patients, showed that the Prevotella/Bacteroides ratio was higher in anti-PD-1 responders (Peng et al., 2020). Anti-PD-1 responders showed an increased abundance of Lachnoclostridium, Parabacteroides, Lachnospiraceae, Ruminococcaceae, Flavonifractor and Dialister, whereas non-responders showed an increased abundance of Bacteroides, Parabacteroides, Coprococcus and Subdoligranulum. SCFA-producing bacteria, such as Eubacterium, Lactobacillus and Streptococcus were associated with better anti-PD-1 efficacy for GI cancers in general.
Table 3 Bacteria and their correlation with ICB efficacy in CRC mice and GI cancer patients.
In CRC mice, it was shown that treatment with antibiotics and anti-PD-1 resulted in higher tumor volumes, indicating that a homeostatic gut microbiome is needed to achieve an optimal anti-PD-1 response (Xu et al., 2020). Furthermore, Prevotella sp. CAG:1031 and Akkermansia muciniphila were related to a better anti-PD-1 response, whereas Bacteroides and Bacteroides sp. CAG:927 appeared to be related to a poorer anti-PD-1 response. A significant upregulation of the glycerophospholipid metabolism could be observed in anti-PD-1 responders. This led the authors to speculate that differences in the microbiome affect the glycerophospholipid metabolism, which alters IFN-γ and IL-2 expression in the tumor microenvironment, resulting in better anti-PD-1 response. When fecal samples from healthy volunteers were administered to GF mice, the extent of IFN-γ+ CD8+ T-cell induction was donor dependent (Tanoue et al., 2019). Eleven species were positively associated with IFN-γ+ CD8+ T-cell induction: Parabacteroides distasonis, Parabacteroides gordonii, Alistipes senegalensis, Parabacteroides johnsonii, Paraprevotella xylaniphila, Bacteroides dorei, Bacteroides uniformis JCM 5828, Eubacterium limosum, Ruminococcaceae bacterium cv2, Phascolarctobacterium faecium and Fusobacterium ulcerans. Furthermore, MC38-tumor bearing mice showed a better response to anti-PD-1 and anti-CTLA-4 treatment after engraftment with these eleven strains. Treatment of the more physiological AOM/DSS CRC mouse model with anti-CTLA-4 or anti-PD-L1 led to smaller and fewer tumors, reduced cancer stem cells, increased immune cell infiltration into tumors, increased CD8+ T-cell frequencies in tumor draining lymph nodes and increased splenic CD4+ and CD8+ T-cell activation (Mager et al., 2020). The metabolite inosine seemed to be responsible for enhancing ICB efficacy. At microbiome level, Bifidobacterium pseudolongum, Olsenella sp., Colidextribacter sp., Bacillus thermoamylovorans, Prevotella sp., Lactobacillus reuteri, Akkermansia muciniphila could only be found in ICB-treated tumors, whereas Collinsella sp., Clostridium cocleatum and Bacteroides sp. were exclusively found in control-treated tumors. Afterwards, it was shown that Bifidobacterium pseudolongum and Olsenella sp. were associated with better ICB-efficacy. Another study on AOM/DSS CRC mice discovered that treatment with anti-CTLA-4 and lysates of Lactobacillus acidophilus reduced the amount of tumors and Tregs and M2 macrophages in mesenteric lymph nodes and increased the amount of IL-2 and IFN-γ in the serum, and CD8+ T-cell infiltration in the tumor (Zhuo et al., 2019). Furthermore, combined treatment with anti-CTLA-4 and Lacobacillus acidophilus lysates restored the dysregulated CRC microbiome by reducing the abundance of Proteobacteria that was increased after tumor development.
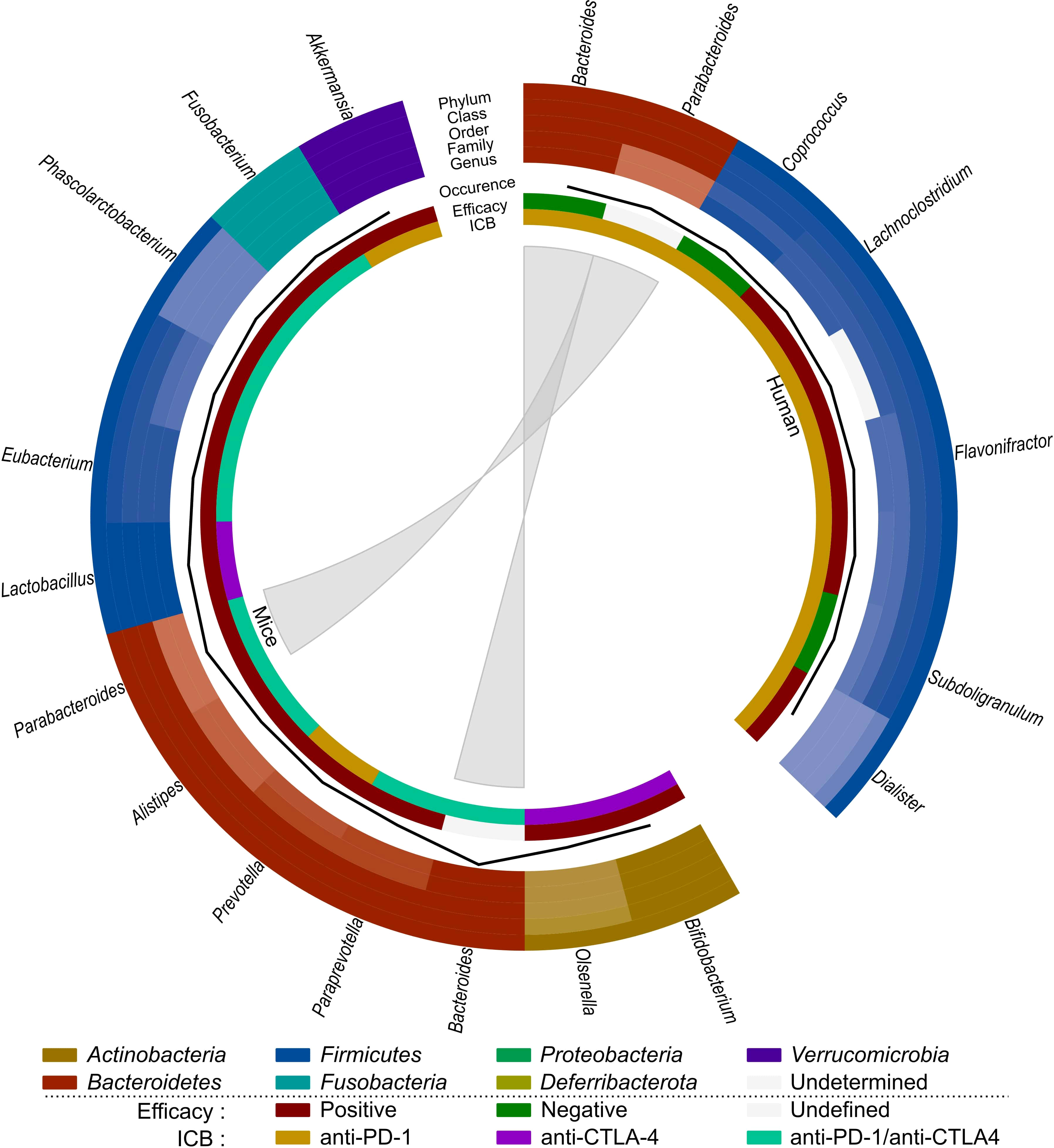
Based on these observations (Table 3; Figure 4), SCFA-producing bacteria, such as Lactobacillus and Eubacterium, seem to enhance immunotherapy efficacy in both mice and humans. Parabacteroides and Akkermansia also seem to be associated with better immunotherapy efficacy in mice, which contradicts their previously mentioned role in radiation-induced toxicity. Bacteroides seems to be negatively related to immunotherapy efficacy in humans, but its impact on immunotherapeutic efficacy in mice is unclear and requires further investigation. As Coprococcus is associated with radiation-induced toxicity as well as worse ICB efficacy, it is a potential detrimental genus for both treatments. Finally, since the role of the gut microbiome on immune-related adverse events has not yet been investigated for CRC, this is an interesting subject for prospective research.
Figure 4 Comparison of the effect of immunotherapy on the gut microbiome of mice and humans. Each distinct phylum is represented by a separate color, as are distinct taxonomic classes, orders, families, and genera. Similar observations are connected. Additional information is provided for each observation, including its occurrence in literature (line, scale maximum is 4), impact on efficacy (increased: red; decreased: green; undefined: grey) and ICB studied (anti-PD-1: yellow; anti-CTLA-4: purple; anti-PD-1/anti-CTLA-4: teal).
5 Combined radio- and immunotherapy treatmentThe view of radiotherapy as a simple cytotoxic agent has dramatically changed in recent years. It is now accepted that radiotherapy can reshape the tumor microenvironment by modulating the immune response (Frey et al., 2017). Therefore, there is a rationale to use immunotherapy together with radiotherapy to boost therapeutic outcomes. However, there is still a need to define biomarkers to identify patients who would benefit most from dual radio- and immunotherapy, optimize optimal sequences/schedules for combined radio- and immunotherapy and identify mechanisms to overcome resistance (Mondini et al., 2020). Furthermore, the effects of dual radio- and immunotherapy on healthy tissues and related toxicity remain largely unknown.
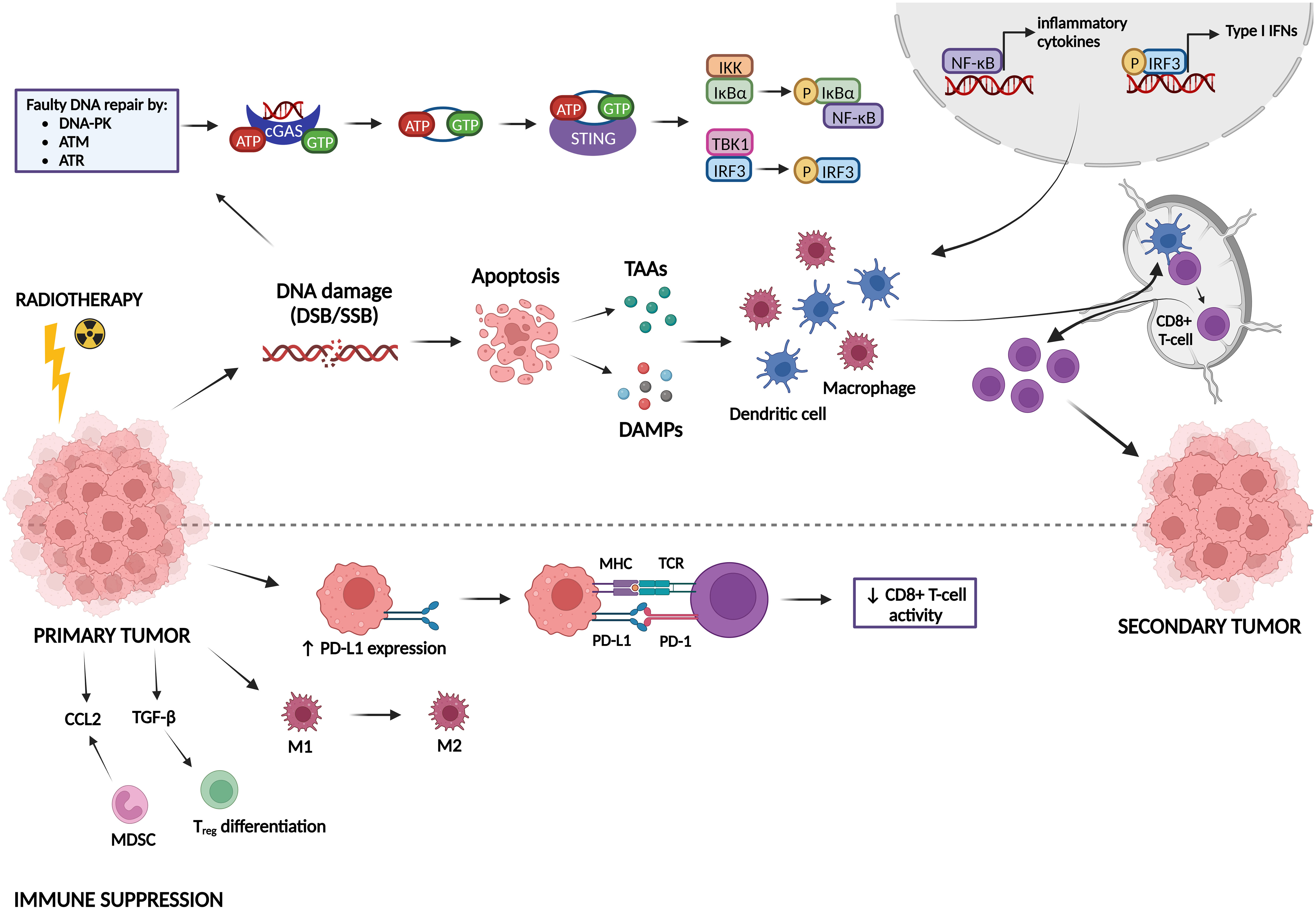
5.1 Immune activating and suppressing effects of radiotherapyRadiotherapy can induce both immune-activating and immune-suppressing effects, which are summarized in Figure 5. Radiotherapy can promote cancer cell killing by inducing DNA damage, but can also activate the immune system, which can be observed by the induction of the abscopal effect and immunogenic cell death (ICD) (Golden and Apetoh, 2015; Brix et al., 2017; Wang et al., 2018). The abscopal effect, which is defined as tumor responses at sites distant from the irradiated site, was first observed by Dr. R. H. Mole in 1953 (Mole, 1953). However, abscopal effects after radiotherapy are rarely observed in the clinic because it is hard to induce an immune response in non-irradiated metastases that are characterized by a specific microenvironment and because metastases might be antigenically heterogeneous (Vanpouille-Box et al., 2018). Radiotherapy can induce the abscopal effect by activation of CD8+ T-cells by antigen-presenting cells (APCs) that have captured radiation-released tumor-associated antigens (TAAs) (Baba et al., 2020; Buchwald et al., 2020). ICD is a type of cell death that promotes a T-cell mediated immune response against antigens derived from dying cells (Kroemer et al., 2013). Radiotherapy induces cell death which generates the release of damage-associated patterns (DAMPs), eventually resulting in ICD (Keam et al., 2020). There are three major DAMPs that contribute to ICD: calreticulin (CRT), high-mobility group box-1 (HMGB-1) and adenosine triphosphate (ATP). CRT can be found on the outer leaflet of dying tumor cells as an “eat me” signal for APCs, leading to presentation of TAAs to naïve T-cells which will result in an anti-tumor immune response (Gameiro et al., 2014). HMGB-1 is released from irradiated tumor cells into the immune environment where it stimulates DCs and macrophages to transcribe inflammatory genes (Wang et al., 2018). ATP is known as a “find me” signal for DCs and monocytes by binding to their purinergic P2X7 receptors, leading to the release of cytokines, such as IL-18 and IL-1β (Perregaux et al., 2000; Ghiringhelli et al., 2009; Aymeric et al., 2010). Radiotherapy can also induce anti-tumor immunity via the cGAS-STING pathway. Radiotherapy-induced DNA damage can be repaired by three central DDR kinases: DNA-dependent protein kinase (DNA-PK), ataxia telangiectasia-mutated (ATM) and ataxia telangiectasia & Rad3-related protein (ATR) (Zhang et al., 2022). These kinases potentiate two main repair mechanisms: non-homologous end-joining and homology-directed repair (Zhang et al., 2022). However, cancer cells often make mistakes during this repair process, which causes genomic instability and cell cycle checkpoint disruption eventually leading to the formation of DNA-containing micronuclei in the cytoplasm (Jeggo et al., 2016; Gekara, 2017). This DNA is then recognized by cyclic GMP-AMP synthase (cGAS) and is dimerized into a cGAS-DNA complex to catalyze the formation of cyclic guanosine monophosphate-adenosine monophosphate (cGAMP). cGAMP interacts with stimulator of IFN genes (STING) and activates it. STING then recruits tank-binding kinase-1 (TBK1) and IκB kinase (IKK) to phosphorylate interferon regulatory factor 3 (IRF3) and NF-κB inhibitor IκBα, respectively. Afterwards, IRF3 and NF-κB are translocated into the nucleus to induce the transcription of type I IFN genes and other inflammatory cytokines like IFN-β (Cai et al., 2014; McLaughlin et al., 2020; Xu et al., 2021). Type I IFN genes facilitate DC maturation, increase DC co-stimulatory molecule expression and enhance DC lymph node migration, which all induces CD8+ T-cell priming (Zitvogel et al., 2015; Sprooten et al., 2019).
Figure 5 Immune activating and suppressing effects of radiotherapy. Radiotherapy induces DNA damage, which can lead either to DNA repair or to apoptosis. Faulty repaired DNA stimulates the cyclic GMP-AMP synthase (cGAS)-stimulator of IFN genes (STING) pathway, leading to the transcription of type I IFN genes and other inflammatory cytokines. In addition, apoptotic cells release tumor-associated antigens (TAAs) and damage-associated molecular patterns (DAMPs). All these factors can activate dendritic cells (DCs) and macrophages, which stimulate CD8+ T-cell priming, inducing immunogenic responses against a secondary tumor. On the other hand, radiotherapy can also act in an immunosuppressive manner. It induces upregulation of PD-L1 expression on tumor cells, leading to inhibition of CD8+ T-cell activation. Furthermore, radiotherapy can promote differentiation/attraction of immunosuppressive immune cells.
Next to all the immune-activating effects of radiotherapy, it has also been shown to induce immunosuppression (Zhang et al., 2022). In this sense, radiotherapy can upregulate the expression of the immune checkpoint PD-L1, thereby inhibiting T-cell activation (Deng et al., 2014; Twyman-Saint Victor et al., 2015). Furthermore, radiotherapy may contribute to resistance mechanisms by reshaping immune cells in the tumor microenvironment towards an immunosuppressive phenotype (Dar et al., 2022). Whereas low-dose radiation can switch tumor-associated macrophages (TAMs) towards the M1 phenotype (pro-inflammatory/anti-tumorigenic), high-dose radiation promotes the M2 phenotype (anti-inflammatory/pro-tumorigenic) (Klug et al., 2013; Prakash et al., 2016; Seifert et al., 2016). Radiotherapy also induces an upregulation of a specific type of immunosuppressive cells called myeloid-derived suppressor cells (MDSCs) and Tregs in the tumor microenvironment (Xu et al., 2013; Muroyama et al., 2017). TGF-β and CCL2 are also upregulated after radiotherapy, resulting in Treg differentiation and attraction of MDSCs to the tumor microenvironment, respectively (Vanpouille-Box et al., 2015; Kalbasi et al., 2017; Dahmani and Delisle, 2018).
5.2 The influence of radio- and immunotherapy on each otherThe abscopal effect of radiotherapy is not commonly observed but has been shown to be synergistically boosted by immunotherapy (Ngwa et al., 2018). Multiple preclinical and some clinical studies have shown an abscopal effect induced by dual radio- and immunotherapy (Demaria et al., 2005; Dewan et al., 2009; Yasuda et al., 2011; Deng et al., 2014; Grimaldi et al., 2014; Golden et al., 2015; Twyman-Saint Victor et al., 2015; Habets et al., 2016; Hao et al., 2016; Theurich et al., 2016; Young et al., 2016; Aboudaram et al., 2017; Dovedi et al., 2017; Koller et al., 2017; Rodriguez-Ruiz et al., 2017; Formenti et al., 2018; Rodríguez-Ruiz et al., 2018; Roger et al., 2018). However, there is a discrepancy in observed abscopal effects induced by dual radio- and immunotherapy between preclinical and clinical studies. It seems that preclinical studies show more promising results than clinical studies, which could be because murine models are not fully able to recapitulate metastatic cancer in patients. Mice are frequently injected with conventional cell lines, which lack genetic and environmental heterogeneity, and typically exhibit only minimal tumor growth (Olson et al., 2018; Arina et al., 2020). More recently, organoid mouse models of CRC have been established for testing radiotherapy (Kim et al., 2022; Nicolas et al., 2022b; Nicolas et al., 2022a). However, these models have yet to be utilized for immunotherapy testing and microbiome research. Radiotherapy and immunotherapy have been shown to enhance each other’s effects. Radiotherapy can enhance immunotherapy efficacy by reducing tumor burden, changing the tumor microenvironment and increasing T-cell infiltration into the tumor (Arina et al., 2020). Furthermore, most patients receiving immunotherapy need additional interventions to overcome primary or acquired resistance to immunotherapy (Vanpouille-Box et al., 2018). Radiotherapy can help to overcome at least some of the mechanisms by which cancer cells are or become resistant to immunotherapy. Radiotherapy generates T-cells specific for TAAs by inducing ICD. It overcomes T-cell exclusion from the tumor by promoting the release of chemokines that attract effector T-cells. It improves recognition and killing of cancer cells by CD8+ T-cells by promoting antigen presentation on MHC class I molecules, thereby upregulating death receptors and promoting the exposure of NK cell-activating ligands (Vanpouille-Box et al., 2018). On the other hand, immunotherapy positively influences radiotherapy by boosting radiotherapy-induced immune activation, blocking immunosuppressive effects of radiotherapy such as PD-L1 upregulation and eliminating microscopic tumors (Arina et al., 2020).
A couple of clinical trials investigating the effect of dual radio- and immunotherapy in CRC patients have been performed. These trials showed little success in shrinking non-irradiated tumors, but did show that the combination treatment was safe and induced an anti-tumor immune response (Monjazeb et al., 2021; Parikh et al., 2021; Segal et al., 2021). Currently, there are even more ongoing clinical trials investigating the effect of dual radio- and immunotherapy in CRC patients (NCT02437071, NCT02837263, NCT04575922, NCT03104439, NCT03101475, NCT02888743).
When MC38-tumor bearing mice were treated with irradiation (20 Gy) and anti-PD-L1 (four injections of 200 µg spread over 72 hours), the abscopal effect could be observed, leading to a significant decrease in tumor volume compared to monotherapy (Deng et al., 2014). In addition, dual radio- and immunotherapy activated CD8+ T-cells, which induced a reduction in MDSCs via TNF-mediated apoptosis, leading to more efficient tumor regression. Treatment of CT26-tumor bearing mice with radiotherapy (5 x 2 Gy) and anti-PD-1 or anti-PD-L1 (10 mg/kg, 3 injections/week for 3 weeks) was curative in 66% and 80% of mice, respectively (Dovedi et al., 2014). It was discovered that radiotherapy activates CD8+ T-cells, which produce IFN-γ, eventually leading to an upregulation of PD-L1 expression on tumor cells. Therefore, the efficacy of the combination treatment can be explained by the fact that immunotherapy blocks the radiotherapy-induced upregulation of PD-L1, allowing a better anti-tumor immune response. Furthermore, when long-term surviving mice (i.e., mice that completely rejected tumor after combination treatment) were rechallenged with tumor cells, they were able to reject the tumor again, indicating that the combination of radio- and immunotherapy generates protective immunologic memory. Another study by the same group investigating the effect of dual radio- and immunotherapy in mice with two tumors, of which only one was irradiated, showed that dual treatment was able to generate anti-tumor responses and tumor control in both irradiated and non-irradiated tumors (Dovedi et al., 2017). It was found that radiotherapy is able to induce polyclonal T-cell infiltration and expansion at the site of treatment, but not in the out-of-field tumor due to suppression through the PD-1/PD-L1 axis. Inhibition of this axis led to a polyclonal T-cell response capable of mediating out-of-field effects. When different radiation schemes (18 x 2 Gy, 3 x 8 Gy or 1 x 16.4 Gy) were applied on CT26-tumor bearing mice in combination with anti-PD-L1 treatment, the best tumor control and survival was observed for the 18 x 2 Gy scheme (Grapin et al., 2019). This indicates that hyperfractionation appears to be critical for lymphoid stimulation, while normo-fractionation seems to be deleterious to lymphoid cells, which are radiosensitive.
It is clear that preclinical studies investigating dual radio- and immunotherapy for CRC are hopeful, whereas clinical studies are not yet providing the same hopeful results. Furthermore, there is no information yet available about the influence of the gut microbiome for this combined treatment of CRC, making this an interesting topic to investigate further.
6 Modulating the gut microbiome to enhance therapeutic modalities for CRCThe gut microbiome can be altered in different ways (Rebersek, 2021). First, biotic interventions, such as diet (e.g., protein, fat, fiber intake) and vitamin intake can change its composition (Singh et al., 2017; Leeming et al., 2019; Pham et al., 2021). In addition, the gut microbiome can also be beneficially altered by pro-, pre- and postbiotics, which are microorganisms, substrates selectively utilized by host microorganisms, and preparations of inanimate microorganisms and/or their components, respectively (Hill et al., 2014; Gibson et al., 2017; Chen et al., 2021; Salminen et al., 2021). A subset of these pro- and prebiotics are designated as psychobiotics, which can boost the mental health status through modulation of the gut microbiome, and as such affect the gut-brain axis (Sarkar et al., 2016). A significant proportion of cancer patients suffer from mental health problems, such as anxiety and depression, which seems to have a negative influence on the gut microbiome (Clapp et al., 2017; Vucic et al., 2021). In the end, this can result in negative treatment outcomes, such as a higher incidence of side effects or less therapy response. Therefore, for these patients, psychobiotics can help to stimulate SCFA production in the gut, leading to the production of gut hormones that migrate to the central nervous system. This cascade induces the release of neurotransmitters, such as dopamine and serotonin, leading to a general decrease in inflammation and restoration of the gut barrier. Another way to modulate the microbiome is by using selective antibiotics that inhibit detrimental bacteria, increase the abundance of certain bacteria that promote cancer therapeutic efficacy or reduce treatment-related side effects by indirectly inhibiting detrimental bacterial metabolites (Wallace et al., 2010; Bullman et al., 2017; Zitvogel et al., 2017). Lastly, fecal microbiota transplantation (FMT), which is the administration of fecal material from a donor into the intestinal tract of a recipient to alter the host’s gut microbiome composition, can be used for therapeutic benefit (Wang et al., 2019). There are multiple methods to deliver FMT, yet the most optimal delivery route remains unclear (Ramai et al., 2019). FMT can for instance be delivered via the upper (via esophagogastroduodenoscopy (EGD), nasogastric, nasojejunal, nasoduodenal tubes or oral capsules) or lower (colonoscopy, sigmoidoscopy and enema) GI route (Gulati et al., 2020). Furthermore, FMT can either be autologous (a-FMT), defined as transplantation of one’s own fecal material taken during healthy state, or heterologous (h-FMT), defined as transplantation of fecal material from a healthy donor to a diseased host (Basson et al., 2020). FMT has been shown to be an effective treatment for recurrent Clostridium difficile infection, which led to its approval by the FDA for this disease (Kelly et al., 2014; Agrawal et al., 2016). Currently, this is the only indication to use FMT in patients. However, the potential beneficial role of FMT for other diseases, such as IBD, functional bowel disorders, metabolic syndrome, autism and multiple sclerosis, is under investigation in clinical trials (Cui et al., 2015; Tian et al., 2016; Paramsothy et al., 2017; Li et al., 2021; Al et al., 2022). Furthermore, FMT has been shown to a have a direct beneficial effect on CRC by reducing inflammation and reducing the abundance of cancer-promoting bacteria (Chen et al., 2019; Kaźmierczak-Siedlecka et al., 2020). Given the importance of FMT, its role in radio- and immunotherapy for CRC is discussed in more detail hereunder.
7 FMT to counteract radiotherapy-related side effectsRadiation induces dysbiosis and GI side effects, which might be reverted by FMT. The use of FMT to reduce dysbiosis and GI side effects has therefore been investigated. A clinical trial investigating the use of FMT as a treatment for chronic RE after abdominal/pelvic irradiation has been performed in five female patients with gynecological cancer (Ding et al., 2020). FMT from healthy donors (18-24 years) was administered for a maximum of three times over a maximum of two weeks through nasojejunal transendoscopic enteral tubing (TET). FMT led to amelioration in rectal hemorrhage, fecal incontinence, diarrhea and abdominal and rectal pain in three out of five recruited patients. However, the efficacy of FMT was not long lasting, indicating that patients may require repeated cycles of FMT. Another case report of a 59-year-old woman with chronic RE after pelvic irradiation showed that FMT from her 18-year-old son improved her chronic RE-related symptoms (Liu et al., 2022). For radiation proctitis after pelvic irradiation, a case report of a 45-year-old woman has shown that four courses of FMT from healthy donors (21 – 24 years) ameliorated the patient’s hematochezia, abdominal pain and diarrhea (Zheng et al., 2020). Furthermore, FMT decreased the abundance of the Firmicutes phylum and increased the abundance of the Bacteroidetes phylum. In the previously mentioned studies, FMT was able to increase the diversity of the patient’s microbiome and shift its composition to one similar of that of the donor (Ding et al., 2020; Zheng et al., 2020).
In another study, mice received fresh FMT from healthy, age-matched mice every day for 10 days via oral gavage after radiation treatment (Cui et al., 2017). They discovered that FMT increased the survival rate and body weight, improved GI tract function and epithelial integrity and enhanced angiogenesis without accelerating tumor growth. FMT was also able to restore the gut microbiome, since it increased the abundance of Bacteroides, Lactobacillus and Prevotella, which were decreased after irradiation. Differences between male and female mice could also be observed, in that male mice that received FMT had increased expression of genes involved in the innate and adaptive immune system, whereas female mice showed increased expression of genes involved in metabolism. In follow-up studies, it was discovered that the abundance of the metabolite indole 3-propion
留言 (0)