
記住我
Gamma-aminobutyric acid type B receptors (GABABRs) are G-protein coupled receptors (GPCR) for GABA, which together with glycine represent the major inhibitory transmitters in the mammalian nervous system. They are widely distributed in nervous tissue, where they regulate neuronal excitability, oscillatory activity and neurogenesis, and are involved in processes such as synaptic plasticity, memory formation and nociception (reviewed in (1–7). Alterations in GABAB functions have been linked to a variety of neurological states and psychiatric disorders including drug addiction, anxiety, cerebral ischemia, depression, epilepsy, neuropathic pain, and spasticity, and Alzheimer’s disease (7–10). In the auditory system, GABABR subunit expression generally shows high levels, particularly in the cochlea, cochlear nucleus, inferior colliculus, medial geniculate nucleus and auditory cortex (11–18). Increasing evidence suggests their functional involvement in the neural circuits that make up these areas, and it is emerging that dysfunction of GABABRs could play a role in some pathologies of the auditory system. In this review, we summarize the current knowledge on the distribution and functions of GABABRs in the peripheral and subcortical parts of the central auditory system obtained in animal studies, mostly using various rodent models.
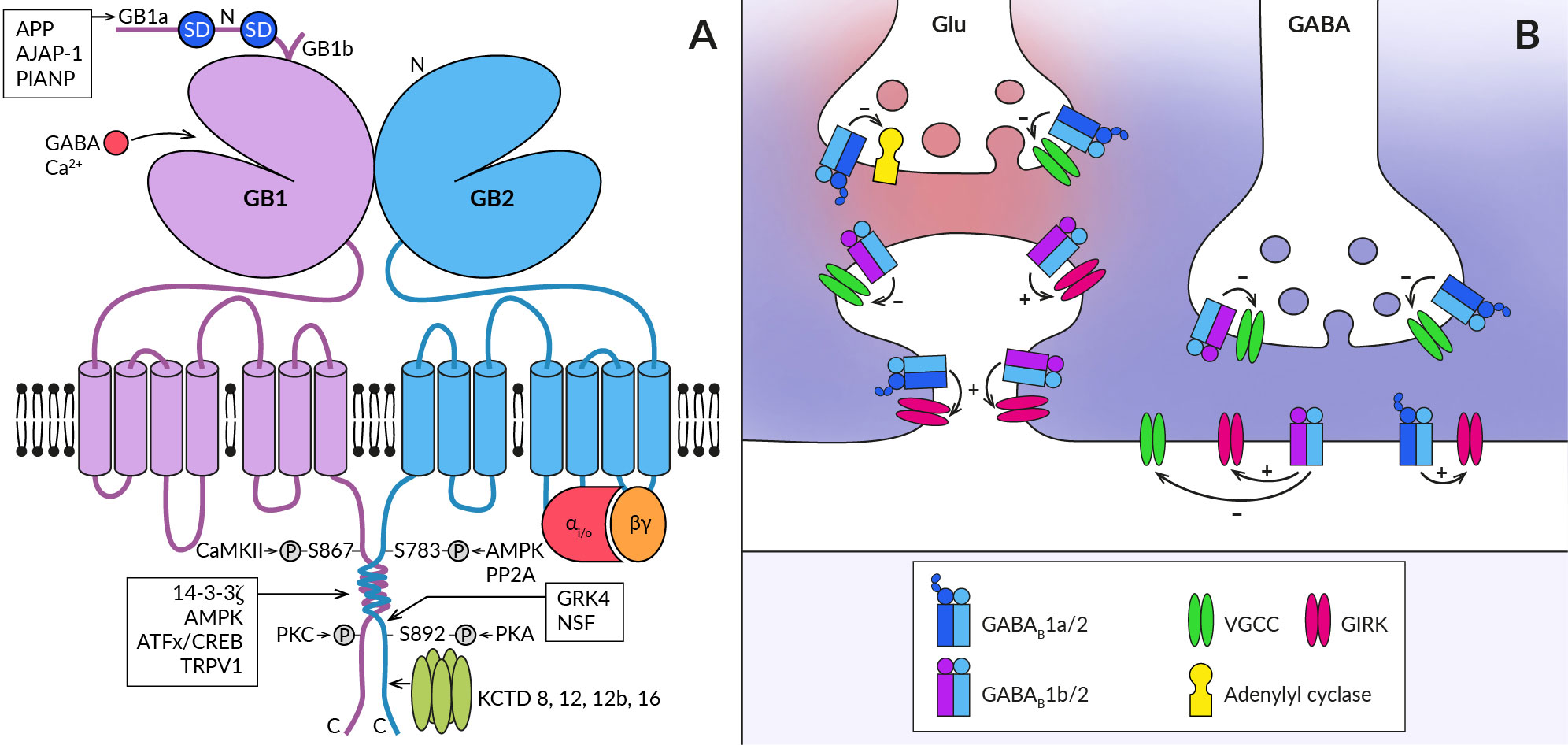
2 Structural basis for heterogeneity in GABABR functionsGABABRs are obligatory heteromers of GABAB1 and GABAB2 subunits (19) (Figure 1A). The subunits heterodimerize through a C-terminal coiled-coil domain which displaces an ER retention signal protein from GABAB1, thus allowing expression of the assembled complexes on the plasma membrane (20, 21). Despite their structural similarity, the subunits play different functional roles in the activation of the receptor heteromer. During this process, GABAB1 mediates agonist binding to GABABR through its N-terminal ‘flytrap’ domain. This induces a series of conformational changes of the receptor and activation of the Gi/o protein via GABAB2 intracellular loops (for a review, see (22–24). Upon activation, the G protein Gα subunits inhibit adenylyl cyclase to decrease cytosolic cAMP levels while the Gβγ subunits inhibit voltage-gated Ca2+ channels (VGCC) or open inwardly rectifying Kir3 K+ channels (GIRK) (25–27). Through coupling to these effector enzymes and ion channels, GABABRs act as important regulators of neurotransmitter release, neuronal excitability and propagation of dendritic spikes (28–32) but see (33).
Figure 1 Schematic representation of the GABABR heterodimer and coupling of its subtypes to effectors at central synapses. (A) The functional GABABR consists of two subunits, GABAB1 (GB1) and GABAB2 (GB2). Both subunits contain large extracellular N-terminal domains (N), seven transmembrane domains connected by three intracellular and three extracellular loops, and an intracellular C-terminus. The N-terminal domain of GABAB1 contains a binding site for the agonist (GABA) and for the endogenous positive allosteric modulator (Ca2+). The two most common splice variants of GABAB1 (GB1a and GB1b) differ in the presence of two sushi domains (SD) at the N-terminus of GABAB1a. Sushi domain binding proteins, the β-amyloid precursor protein (APP), the adherence junction-associated protein 1 (AJAP-1), and the PILRα-associated neural protein (PIANP) form complexes with GABAB1a/2 receptors. The GABAB2 subunit interacts with heterotrimeric G-proteins (αi/o, βγ) and stimulates their activation. At least four phosphorylation sites were identified on GABABR subunits: S867 and a yet unidentified site at the C-terminus on the GABAB1 subunit are phosphorylated by calcium/calmodulin-dependent protein kinase II (CaMKII) and protein kinase C (PKC), respectively; on the GABAB2 subunit, S783 is phosphorylated by 5′AMP-dependent protein kinase (AMPK) and S892 is phosphorylated by protein kinase A and dephosphorylated by protein phosphatase 2A (PP2A). Other GABAB1 interacting proteins include 14-3-3 proteins, the capsaicin receptor TRPV1 and ATF/CREB family transcription factors. By specifically interacting with the latter, GABABR can directly influence gene expression. GABAB2 can further associate with G-protein receptor kinase 4 (GRK4) and N-ethylmaleimide-sensitive factor (NSF), leading to the regulation of GABABR activity. The C-terminus of GABAB2 contains a binding site for auxiliary receptor subunits, proteins of the potassium channel tetramerization domain (KCTD) family. (B) GABABRs are expressed in presynaptic and postsynaptic compartments of both excitatory (Glu) and inhibitory (GABA) synapses (synaptic glutamate and GABA are shown in red and blue, respectively). They associate with effector enzymes and ion channels (adenylyl cyclase, VGCC, GIRK) to regulate neurotransmitter release and neuronal excitability. GABAB1a-containing receptors (GABAB1a/2) are preferentially localized in the presynaptic membrane of both types of synapses and less frequently at postsynaptic sites such as the dendritic shaft or spine neck. In contrast, GABAB1b-containing receptors (GABAB1b/2) prefer postsynaptic sites but are also expressed in inhibitory terminals. GABABRs at inhibitory synapses are activated by synaptic GABA and can mediate slow inhibitory postsynaptic currents. Heteroreceptors at excitatory synapses are activated either tonically by ambient agonist concentration or require GABA spillover from neighboring inhibitory synapses.
The physiological functions of GABABRs critically depend on their density and location in specific neural compartments, as well as on the kinetics of their signaling. These properties of GABABR are, on the other hand, significantly modulated by its post-translational modifications and interactions with associated proteins. First, ample evidence exists that phosphorylation of GABABR subunits by serine/threonine protein kinases bidirectionally affects both cell surface receptor expression and the magnitude of its responses (for details, see (34, 35). Phosphorylation of GABAB1 at S867 by calcium/calmodulin-dependent kinase II or at an unidentified site by protein kinase C triggers GABABR internalization or desensitization (36, 37). Conversely, phosphorylation of GABAB2 by adenosine monophosphate-activated protein kinase (S783) or cAMP-dependent protein kinase (S892) stabilizes GABABRs at the cell surface (38, 39). Second, the subcellular localization of GABABR and the dynamics of its intracellular trafficking depend, at least in some brain areas, on the type of GABAB1 isoform (Figure 1B). The GABAB1a variant containing two extracellular sushi domains (SDs) at its N-terminus is predominantly expressed in glutamatergic axonal terminals whilst the GABAB1b lacking SDs expresses in somatodendritic parts (31, 40–42). GABAB1a is also present in dendritic shafts but seems to be excluded from the spine heads (40). Recent studies have identified several proteins that interact with SDs (Figure 1A) and promote axonal trafficking of GABAB1a and/or receptor stabilization at the presynaptic plasma membrane (43–45). Among these proteins, the amyloid-precursor protein (APP) appears to play a key role, linking the APP/GABABR complex to the axonal trafficking motor (for details, see (19, 23). Moreover, a secreted cleaved APP fragment has been shown to regulate synaptic transmission in the hippocampus of mice in a GABABR-dependent manner (45), but it appears to act through a more complex mechanism than as a functional receptor ligand (46). GABAB1 subunits have also been reported to associate with 14-3-3 proteins and the capsaicin receptor TRPV1 (47, 48), with both interactions being implicated in chronic pain conditions. 14-3-3 binds to the C-terminus of GABAB1 to dissociate the GABAB heterodimer, resulting in impaired GABAB signaling and reduced control of TRPV1 sensitization in spinal neurons (47–49).
Lastly, four cytosolic K+-channel tetramerization-domain (KCTD) proteins KCTD8, KCTD12, KCTD12b and KCTD16, which constitutively bind to the GABAB2 C-terminal domain as pentamers, increase receptor surface expression and show multiple effects on its response kinetics (50–52). The KCTD proteins comprise the N-terminal T1 and C-terminal H1 domains (53, 54), capable of simultaneous interactions with GABAB2 and Gβγ subunits of the G protein, respectively (55, 56). The preassembled complex is characterized by elevated potency and accelerated kinetics of G-protein signaling (50, 55). In addition, KCTD12 and 12b induce pronounced desensitization of GABABR responses by activity-dependent stripping of Gβγ from GIRK or VGCC channels (55, 56). The desensitization is in turn regulated by phosphorylation of serine-892 on GABAB2 or by heteromerization of KCTD12 with KCTD16 (57). KCTD16 itself slows down the deactivation phase of GABABR activated GIRK currents by unknown mechanism (58). Moreover, KCTD8 and KCTD16 contain a C-terminal H2 domain that binds secondary GABABR interacting proteins, such as VGCC or hyperpolarization-activated cyclic nucleotide-gated channels (43). Because many neurons in the brain simultaneously express several KCTDs and some of them possibly all KCTDs, assembly of distinct KCTDs at the receptor increases the molecular and functional repertoire of native GABABRs (58, 59).
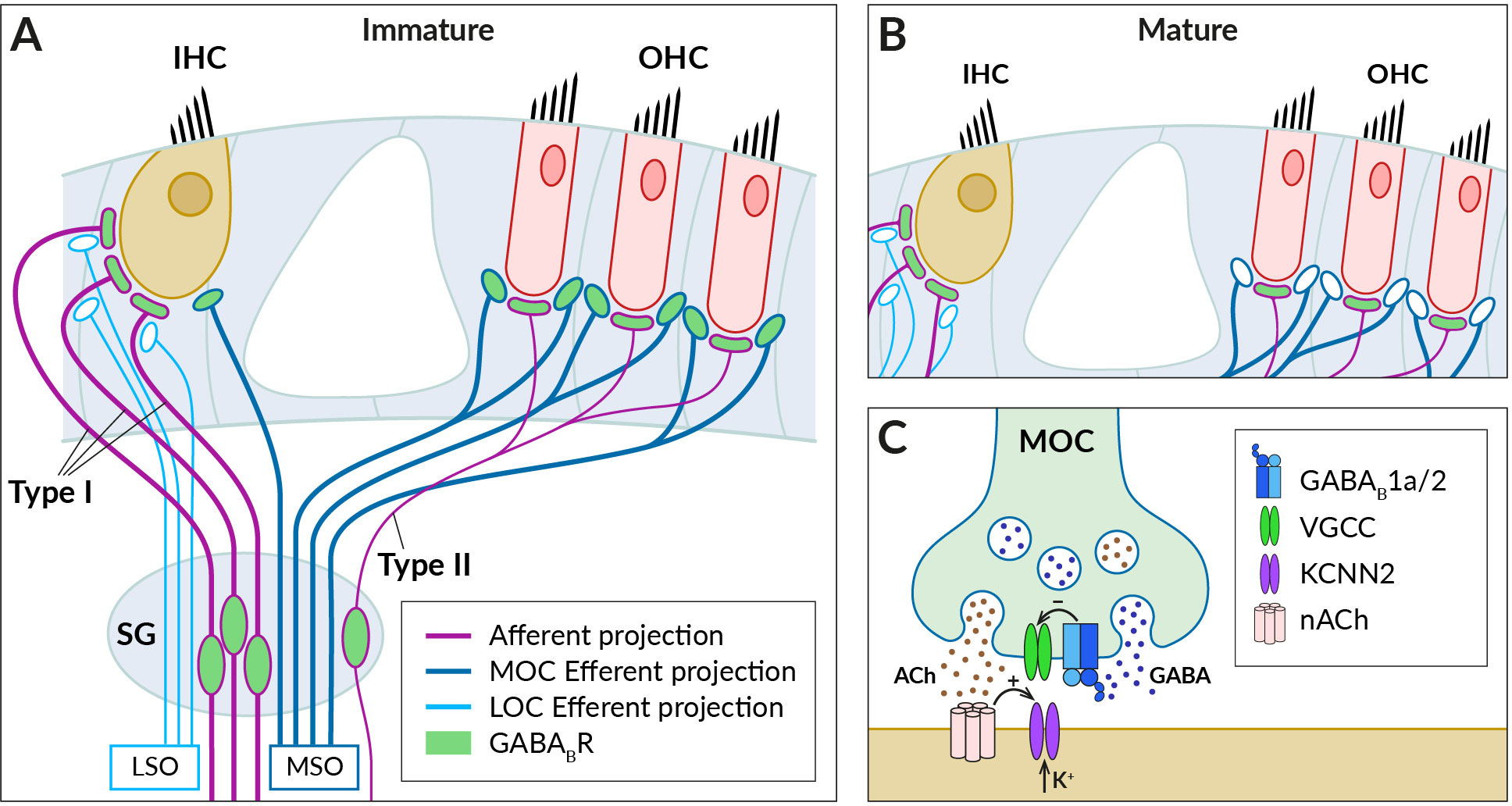
3 Cochlear GABABRsIn the mammalian cochlea, the sensory organ of Corti comprises one row of inner and three rows of outer hair cells (IHCs and OHCs) (Figure 2). Their main tasks are to amplify the incoming auditory signals (OHC) and to relay auditory information to the brain (IHC) (for a review, see (60, 61). Hair cells receive both afferent and efferent innervation. The afferent innervation is carried by spiral ganglion neurons type I and II, which respond to glutamate released by IHCs and OHCs, respectively, and initiate and conduct action potentials to the cochlear nucleus (62–65). GABABRs have been found in both type I and type II spiral ganglion neurons and their afferent terminals at IHCs and OHCs that do not express GABABRs themselves (66–70). Activation of the receptors with baclofen diminished glutamate induced increases of intracellular Ca2+ concentration in cultured spiral ganglion neurons (66). The mechanism underlying the inhibition has not been revealed but these observations would be consistent with GABABR-mediated regulation of the excitability of IHC afferents by reducing the Ca2+ permeability of postsynaptic NMDA receptors (32, 71). In addition, GABABRs could also inhibit Ca2+-dependent K+ conductance in a way found in vestibular hair cell afferents (72–75), thereby modulating the spike frequency in spiral ganglion neurons (73, 76).
Figure 2 GABABRs expressed in fibers innervating cochlear hair cells. (A) Schematic of afferent (purple) and efferent (blue) innervation of inner (IHC) and outer hair cells (OHC) in the immature cochlea showing the known localization of the GABABR (green). Receptors have been found in type I and II spiral ganglion (SG) neurons and their afferent terminals on the IHC and OHC, where they are thought to regulate glutamate-evoked responses, and in the terminals of the medial olivocochlear bundle (MOC), which originates around the medial superior olive (MSO). The presence of GABABRs in fibers of the lateral olivocochlear bundle (LOC) arising from the lateral superior olive (LSO) was not detected. (B) In the adult cochlea, GABABRs disappear from the terminals of efferent fibers, along with efferent innervation of IHC by MOC fibers. (C) Schematic representation of the cholinergic synapse formed by MOC fibers on the IHC somata. At this synapse, GABABRs control the secretion of acetylcholine (ACh, brown dots), which is released together with GABA (blue dots) from the MOC terminal. GABA is thought to activate presynaptic GABAB1a/2 autoreceptors, which regulate acetylcholine release by inhibiting presynaptic VGCC channels. Acetylcholine binds to nicotinic receptors (nACh) on the IHC, which elicit postsynaptic Ca2+ transients and K+ currents via Ca2+-dependent K+ channels (KCNN2) (see section 3 for further details).
The efferent fibers originate from the superior olivary complex and allow feedback control of cochlear activity by the auditory brainstem (for reviews, see (77–79). There are two groups of olivocochlear efferents (OC): medial OC (MOC), large-diameter myelinated fibers arising from regions near the medial superior olive (MSO) and releasing acetylcholine to reduce the gain of the OHC amplifier, and lateral OC (LOC), thinner unmyelinated fibers that arise near the lateral superior olive (LSO) and modulate the excitability of type I afferents by releasing multiple transmitters, including acetylcholine and GABA. Functional GABABRs have been found in OC bundles, where their expression appeared to be developmentally regulated (80) (Figure 2). Mice during the second postnatal week express GABAB1a/2 receptors at axonal terminals of OC fibers innervating somata of OHCs and, at this developmental stage, also IHCs (68) (Figure 2A). The receptors control acetylcholine release at efferent synapses via inhibition of presynaptic P-/Q-type VGCC (68) (Figure 2C). In adult mice, however, no GABABRs or their disinhibitory effects were observed in MOC terminals or OHCs, respectively (67) (Figure 2B).
The role of cochlear GABABRs in the auditory function remains to be understood. The receptors are thought to be activated in vivo by GABA co-released with acetylcholine from LOC and MOC fibers (79–81) (Figure 2C). The firing frequency of the fibers and thus the secretory activity of their efferent synapses depend on the intensity of sound stimulation (77, 82). During low-intensity acoustic stimulation, GABABRs can be activated predominantly by the background GABA (83) and tonically suppress the release of acetylcholine at OC terminals. Consistent with this expectation, selective GABABR antagonists increase the amplitudes of postsynaptic currents elicited in hair cells by low-frequency stimulation of MOC fibers (68). The low basal probability of acetylcholine release then allows MOC-OHC synapses to respond to high-frequency stimulation by strongly facilitating cholinergic postsynaptic currents (78, 84). It has been proposed that the presynaptic facilitation together with a summation of repetitive postsynaptic currents significantly increase the reliability and strength of cholinergic synaptic transmission during high-intensity acoustic stimulation (78). This, in turn, would lead to a greater reduction in the gain of the OHC amplifier when exposed to intense sounds and to the protection of the immature cochlea from acoustic trauma (80). Adult mice with deleted GABABRs exhibit increased hearing thresholds as measured by auditory brainstem responses and distortion product otoacoustic emissions (67). This indicated the importance of GABABRs for the normal function of the mature cochlea. It has been suggested that deletion of GABABRs in spiral ganglion and brainstem neurons led to increased spontaneous activity and elevation of the thresholds (67).
Finally, GABABR-associated KCTD12 proteins have been found to be highly expressed in the mammalian cochlea during the early developmental stages (11, 59). Immunostaining experiments located KCTD12 in spiral ganglion neurons and also in cochlear supporting cells and fibrocytes (11). As these non-neuronal cells have been implicated in the K+ recycling pathways, KCTD12 could play a role in ion transport or ionic content regulation in the cochlea (11, 85, 86).
4 GABABRs in the auditory brainstem and midbrain4.1 The cochlear nucleusThe cochlear nucleus (CN) is the first processing station of the central auditory pathway. It consists of two distinct regions, the dorsal (DCN) and ventral (VCN) cochlear nuclei (Figure 3). Fibers of the auditory nerve in both parts contact the major projection neurons and several types of interneurons to distribute sensory information while maintaining the tonotopic organization. Incoming cochlear signals are then preprocessed by CN circuitry, integrated with signals received by multimodal inputs and conveyed to multiple ascending auditory pathways (88–90). In this way, the CN plays a role in processes such as frequency representation, intensity and time coding, localization of sound sources and filtering of self-generated sounds (91).
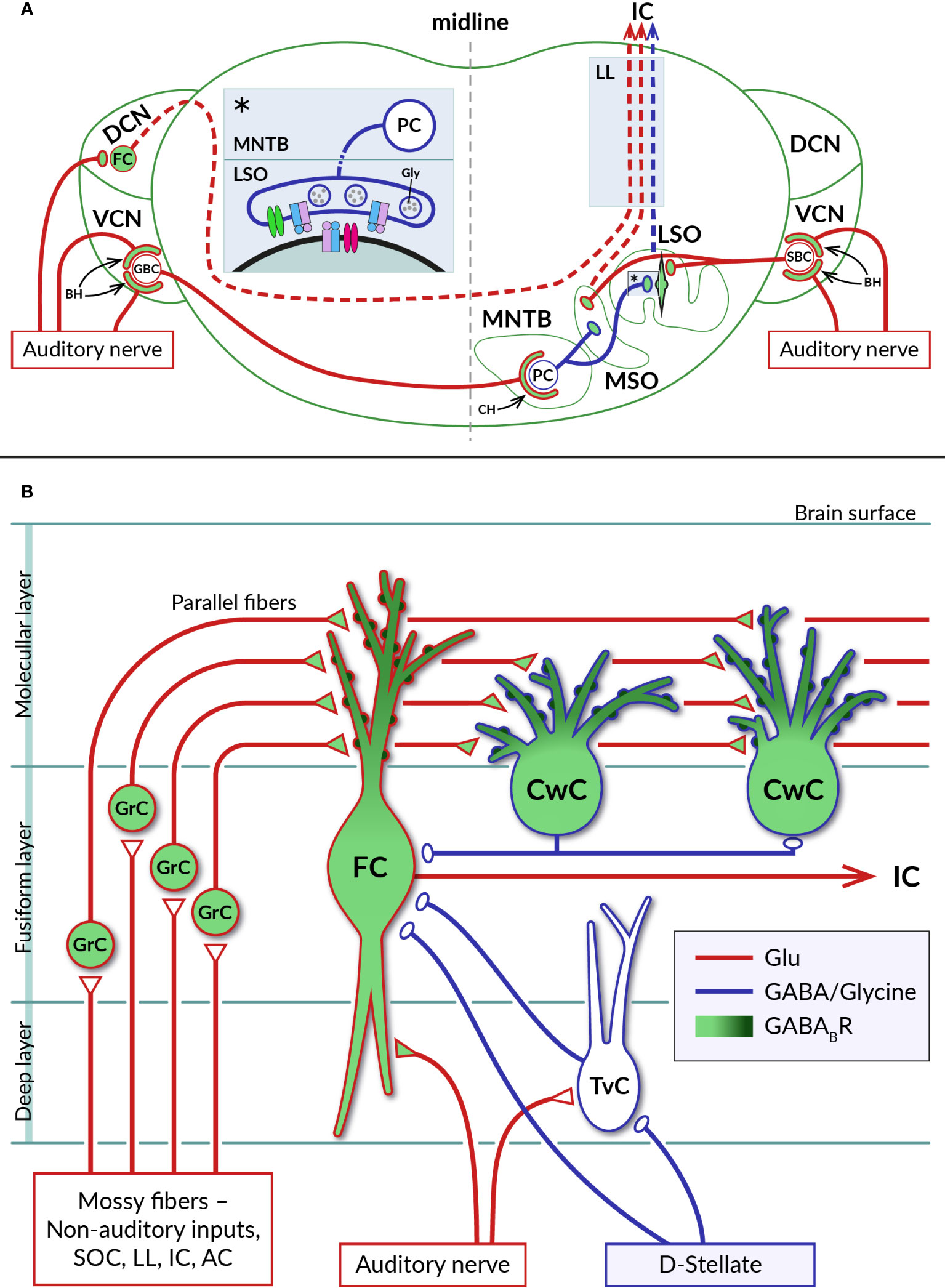
Figure 3 Distribution of GABABRs in auditory brainstem nuclei. (A) Diagram of a coronal section through the brainstem showing a simplified representation of the circuits formed by neurons of the auditory nuclei, dorsal and ventral cochlear nuclei (DCN, VCN), lateral and medial superior olive (LSO, MSO), and medial nucleus of the trapezoid body (MNTB). Excitatory (glutamatergic) and inhibitory (glycinergic) projections are represented by red and blue lines, respectively. GABABRs have been identified on large axosomatic terminals on spherical bushy cells (SBC) and globular bushy cells (GBC) in the VCN and on MNTB principal cells (PC), referred to as endbulbs of Held (BH) and calyces of Held (CH), respectively, as well as on principal neurons in the LSO and MSO and their excitatory and inhibitory boutons. Dashed lines indicate ascending projections of brainstem neurons passing through the lateral lemniscus (LL) to the inferior colliculus (IC). The inset (asterisk) on the left of the section shows a detail of the inhibitory synapse formed by the axon of an MNTB PC on the soma of an LSO neuron. At mature LSO synapses, presynaptic GABABRs control glycine release by inhibiting VGCC (green), whereas at immature synapses, somatic GABABRs additionally regulate postsynaptic excitability by activating GIRK channels (red). See sections 4.1 and 4.2 for details. (B) Schematic representation of selected synaptic connections between excitatory (red) and inhibitory (blue) neurons in the DCN (adapted from (87). GABABRs have been found in pre- and postsynaptic compartments of DCN neurons, where they control glutamate release and short-term synaptic plasticity or neuronal excitability. Presynaptic receptors are localized in auditory nerve endings on basal dendrites of glutamatergic fusiform cells (FC) and in glutamatergic terminals of axons of granule cells (GrC), parallel fibers, innervating apical dendrites of both fusiform cells and glycinergic cartwheel cells (CwC). The subcellular distribution of postsynaptic GABABRs expressed by fusiform and cartwheel cells shows a dendrosomatic gradient, with receptor density reaching highest values in the distal parts of apical dendrites (dark green). AC – auditory cortex, IC – inferior colliculus, LL – lateral lemniscus, SOC – superior olivary complex, TvC – tuberculoventral cell, D-stellate – a subtype of inhibitory neuron in VCN.
The existence of functional GABABRs in the CN was suggested in the pioneering work of Caspary and colleagues (92). Subsequently, GABABRs were found to be involved in CN circuits at pre- and postsynaptic sites, where they control neurotransmitter release, neuronal excitability and short-term plasticity of synaptic currents. Consistent with the expression of GABABRs by spiral ganglion neurons, the receptors have been found at the axonal terminals of the auditory nerve on CN neurons (16, 93) (Figure 3A). In the VCN, type I fibers contact glutamatergic spherical and globular bushy cells, T-stellate neurons, octopus cells, and glycinergic D- and L- stellate interneurons (94–98). GABAB function was mostly studied in the endbulb of Held synapses formed by auditory nerve fibers on somata of bushy cells. Receptor activation has been found to inhibit presynaptic Ca2+ influx through N- or P-/Q-type VGCCs, leading to reduced glutamate release from endbulbs and diminished amplitudes of excitatory postsynaptic potentials in bushy cells (99–101). As a result, GABABR activation reduced the probability of bushy cells to initiate action potentials in response to auditory nerve stimulation. Cell firing could be restored when two converging synaptic inputs were activated simultaneously or by postsynaptic depolarization by group I metabotropic glutamate receptors (99, 100). Thus, in the presence of GABA, bushy cells appeared to function as coincidence detectors with the spiking probability dependent on the synchronous activity of multiple inputs or on modulation via other G-protein-dependent pathways. It has been proposed that the action of presynaptic GABABRs at the endbulb of Held synapses suppresses the relaying of incoming spontaneous activities and enhances the temporal coding observed in bushy cells in vivo (99, 102, 103). Potential sources of endogenous agonists for these receptors could include intrinsic and extrinsic inhibitory synaptic inputs to bushy cells arising from D- or L- stellate cells in the VCN, GABA/glycinergic neurons in the DCN and descending inhibitory projections from the superior olivary complex (97, 104, 105). Accordingly, a pharmacological study has shown that repetitive stimulation of inhibitory synapses formed by D-stellate interneurons on bushy cell bodies leads to GABA accumulation, activation of presynaptic GABAB autoreceptors, and suppression of glycinergic postsynaptic currents (106). However, experimental evidence that GABA, which is released from these synapses along with glycine, also actually activates GABAB heteroreceptors on endbulbs has not yet been obtained.
In the DCN, type I fibers innervate basal dendrites of glutamatergic fusiform cells, giant cells and glycinergic tuberculoventral neurons (107) (Figure 3B). Type II fibers terminate on neurons in the granular cell domain (65, 108) that receive additional inputs from somatosensory and motor systems (109, 110). The axons of granule cells then enter the molecular layer of the DCN and give rise to parallel fibers that excite the spiny apical dendrites of fusiform cells, as well as GABA/glycinergic cartwheel cells, stellate cells and Golgi cells (87). Fusiform cells, the principal DCN projection neurons, thus represent highly integrative units of the ascending auditory system (89). Both types of excitatory synaptic inputs to these cells were found to be controlled by presynaptic GABABRs. Activation of the receptors by baclofen reduced the release of glutamate while decreasing short-term synaptic depression at auditory nerve endings and enhancing the facilitation of release from parallel fibers (93, 111). It has been suggested that in this way presynaptic GABABRs support the sustained transmission of auditory signals to fusiform cells at increased sound intensities and amplify somatosensory information at the parallel fiber synapses formed by high frequency inputs (93). In addition, the excitability of fusiform cells has been found to be regulated by postsynaptic GABABRs coupled to GIRK and N-type VGCC (93, 112). The concentration of these receptors in the cell membrane shows a dendrosomatic gradient, reaching the highest levels in the distal parts of the apical dendrites (15). This indicates that postsynaptic GABABRs could control synaptic inputs from parallel fibers more strongly than those from the auditory nerve.
GABABRs regulating fusiform cell activity are thought to be stimulated by agonists released from inhibitory projections that derive from the superior olivary complex, lateral lemniscus or from local GABA/glycinergic interneurons (Figure 3) (113–115). Of the latter, cartwheel cells are the most abundant inhibitory interneurons in the DCN (116). Their excitation through parallel fiber synapses has been shown to cause a strong inhibition of fusiform cells as well as other cartwheel cells (116–118). They are therefore considered to form the basis for both the feed-forward inhibitory and disinhibitory circuits in the DCN (116, 119, 120). Neuronal processes operating in these circuits likely involve GABABR activity. Cartwheel cells have been found to be highly GABAB1-immunoreactive, similar to fusiform cells (15, 121, 122), although the functional properties of their GABABRs remain unexplored. The receptors could regulate spontaneous activity of cartwheel cells in a way observed for other GPCRs which use the same signaling pathway. Kuo and Trussell (123) found that α2 adrenergic receptors, by eliminating the background spiking of cartwheel cells, relieved their inhibitory synapses from depression, thereby enhancing the stimulus-evoked inhibition of fusiform cells.
4.2 Superior olivary complexThe superior olive complex (SOC) consists of a group of interconnected brainstem nuclei that process binaural information necessary for sound source localization and modulate the function of other auditory areas via the olivocochlear bundle or numerous inhibitory projections (77, 124, 125). Sound-localizing SOC circuits include principal neurons in the medial nucleus of the trapezoid body (MNTB) that convert excitatory signals from the contralateral VCN into properly timed glycinergic inhibition transmitted to the ipsilateral LSO and MSO (126) (Figure 3A). GABABRs have been shown to participate in sound localization mechanisms by modifying the sensitivity of LSO and MSO neurons that encode interaural sound level and time differences (ILDs and ITDs) (see reviews (127, 128), for details). In summary, it has been shown in the gerbil that repeated activation of principal neurons in the LSO and MSO leads to the release of GABA, which then differentially inhibits glutamatergic and glycinergic inputs to these neurons via presynaptic GABABRs (129–131). In both nuclei, GABA is released in an activity-dependent manner, either directly from somatodendritic parts of LSO principal cells or from GABAergic projections to the MSO, allowing feedback control of spiking of LSO and MSO neurons (129, 130). As a result, ILD and ITD responses of neurons show a dependence on their previous spiking activity, suggesting that binaural processing in the SOC is subject to GABABR-mediated dynamic adaptation (128, 132, 133). In addition, as shown in mice and gerbils, GABABRs can be tonically activated by ambient GABA and regulate the excitability of LSO and MSO neurons via postsynaptic GIRK channels (131, 134). These effects are thought to protect principal cells from overexcitation caused by increased spontaneous activity entering the binaural nuclei around the onset of hearing (127). At this stage of auditory system development, GABABRs at glycinergic MNTB-LSO synapses also play important roles in mechanisms of long-term plasticity of inhibitory transmission. Before the onset of hearing, postsynaptic receptors mediate the depression (LTD) of MNTB-evoked inhibitory potentials (135, 136), whereas following the onset, GABABR signaling is required to induce potentiation (LTP) of inhibitory postsynaptic currents (137). Based on these observations, it has been proposed that GABABR-dependent plasticity underlies both the early elimination of redundant inhibitory synaptic connections in the LSO and their later stabilization and strengthening during subsequent postnatal development (137, 138).
Excitatory inputs of MNTB principal cells formed as giant axosomatic terminals, the calyces of Held (139–141) (Figure 3A), allow direct electrophysiological examination (142) and have therefore often been used to study the effects of GABABRs on neurotransmitter exocytosis. It has been found that activation of calyceal GABABRs receptors in rat brainstem slices blocks approximately 80% of glutamate release, by inhibiting presynaptic VGCCs through the action of Gβγ subunits (143–145). In addition, GABABRs can directly interfere with the vesicular cycle by reducing cAMP (30). The endogenous source of agonists for presynaptic GABABRs in MNTB remains unclear. It has been suggested that the receptors could be tonically activated by ambient GABA and subsequently help to maintain the relatively low basal probability of glutamate release and little short-term plasticity observed at the calyx of Held synapse in vivo (146–148). However, in vivo experiments with pharmacological modulation of GABABR activity have not yet confirmed this hypothesis. No significant change in synaptic transmission in mice was observed during application of the GABABR antagonist CGP54626, suggesting a low ambient GABA concentration (149). However, these experiments did not exclude the possibility that the application of antagonist also led to inhibition of presynaptic GABABRs at glycinergic synapses, thereby enhancing inhibitory transmission in the MNTB. This would in turn reduce the intensity of steady-state transmission at the calyx of Held synapse via pre- and postsynaptic glycine receptors, thus compensating for the effect of inhibition of calyceal GABABRs (150–154). Further in vivo experiments using glycine receptor antagonists would be needed to test this possibility.
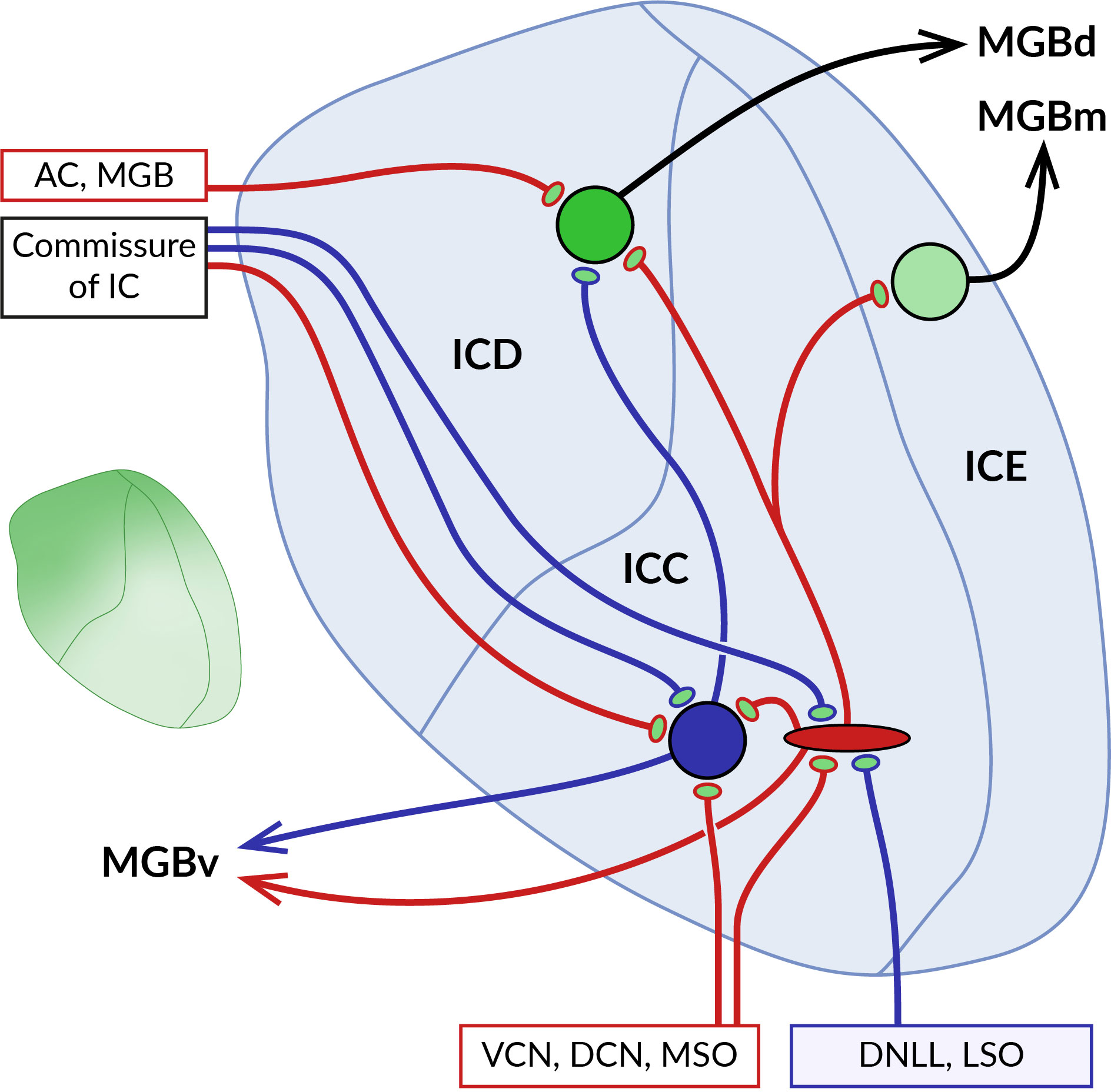
4.3 Inferior colliculusThe inferior colliculus (IC) is a midbrain structure that connects the auditory regions of the hindbrain and forebrain. It consists of a central core surrounded by lateral, dorsal and rostral cortices (125, 155) (Figure 4). Ascending fibers from the auditory brainstem innervate mostly neurons in the central nucleus, while descending fibers from the auditory cortex and thalamus terminate mainly in the external and to a limited extent in the central part of the IC (125, 156). Efferent fibers from the IC ascend to the corresponding parts of the thalamic medial geniculate nucleus or give rise to caudally oriented projections to the lower brainstem (157–159). The two ICs are interconnected by commissural fibers abundantly formed as collaterals of projections of the central core neurons to the ipsilateral medial geniculate body (160). In addition, the IC receives input from numerous non-auditory areas (161). It has been proposed that the function of neurons in the central nucleus is to integrate ascending auditory information and generate de novo coding properties, while those in the IC cortex allow for the integration of multimodal information and the detection of novel stimulus features (162).
Figure 4 Localization of GABABRs at pre- and postsynaptic sites in the inferior colliculus (IC). Schematic representation of the main parts of the IC with their main afferent and efferent projections. The central core of the IC (ICC) is surrounded by the dorsal (ICD) and external cortex (ICE). Ascending excitatory (red) and inhibitory (blue) fibers from ventral and dorsal cochlear nuclei (VCN, DCN), medial superior olive (MSO), dorsal nucleus of the lateral lemniscus (DNLL), and lateral superior olive (LSO) contact large GABAergic and glutamatergic neurons in the ICC. Both types of ICC neurons project to cortical areas, predominantly to the ICD. The two ICs are interconnected by commissural fibers. Descending projections from the auditory cortex (AC) and medial geniculate body (MGB) terminate on ICD neurons. As the diagram shows, neurons in different parts of the IC preferentially project to distinct nuclei of the auditory thalamus, medial, dorsal and ventral MGB (MGBm, MGBd and MGBv). GABABRs in ICC show a preferential presynaptic localization, while cortical receptors occupy both pre- and postsynaptic locations. The inset on the left shows the gradient of the expression level of GABABRs in the IC, which reaches the highest values in the dorsomedial part of the IC and decreases ventrolaterally.
The distribution of GABABRs in the IC was studied in brain sections of rats and big brown bats using quantitative autoradiography and immunohistochemistry (13, 16, 17, 163). The receptors were found throughout the entire IC both in neuronal somata and in the neuropil, with the highest expression levels in the dorsomedial cortical part which was also characterized by an increased density of GABAB-positive cell bodies (see inset in Figure 4). Pharmacological modulation of receptor activity significantly affects the sound-evoked responses of IC neurons (164–168) by a mechanism that appears to differ between IC subdivisions. In the central nucleus, presynaptic GABABRs have been shown to control the release of glutamate and GABA from excitatory and inhibitory fibers that form the lemniscal ascending inputs for sound-driven signals (169–171). The latter GABAB autoreceptors, by suppressing GABAergic inhibition, can also promote the induction of long-term potentiation of excitatory potentials in IC neurons (171). In these studies, no postsynaptic GABABR responses were observed following baclofen application or stimulation of inhibitory fibers, suggesting that the receptors in the central nucleus act primarily at presynaptic sites. Conversely, in the dorsal cortex of the IC, GABABR activation leads to both presynaptic and postsynaptic responses. The former is associated with a reduced release of glutamate and GABA from the endings of afferent fibers, similar to that in the central nucleus (172, 173). Postsynaptic receptor responses, elicited pharmacologically or by stimulation of commissural GABAergic input, involve changes in the activity of GIRK and VGCC effector channels and significantly affect the firing properties of neurons in this region (173). Thus, these data suggest that GABABRs in cortical parts of the IC may modulate sound-evoked neuronal activity through pre- and postsynaptic functions and may also directly mediate inhibitory synaptic transmission.
Endogenous GABABR activation in the IC could be triggered by GABAergic projections originating from the dorsal and ventral nuclei of the lateral lemniscus, the superior paraolivary nucleus and the contralateral IC (155, 174–176), as well as by local inhibitory interneurons. GABAergic neurons account for 25% of all IC neurons (177) and represent a heterogeneous group composed of several subtypes, differing in synaptic organization and neuronal connections (178, 179). A well-studied subset of large GABAergic neurons receives convergent glutamatergic input from multiple sources, including the IC, lateral lemniscus, SOC, DCN, and auditory cortex (180–182), suggesting that these neurons are part of complex feedforward, feedback, or disinhibitory neuronal circuits in the auditory pathway. However, the precise mechanisms by which GABABRs expressed in IC neurons contribute to the function of these circuits have not yet been fully understood. Due to their wide distribution at presynaptic and postsynaptic sites and the relatively slow kinetics of their responses, GABABRs are thought to regulate overall neural sensitivity to sounds and to set the gain of signal processing in the IC (17, 165, 167, 168). This idea would be consistent with the observation that pharmacological receptor blockade increases the acoustic excitation of IC neurons but does not reduce the inhibition elicited by paired stimuli or increase the range of amplitude modulation rates that evoke phase locking (166, 167). Interestingly, antagonists acting at postsynaptic GABABRs were found to reduce response adaptation of specialized IC cortex neurons to repetitive sounds, a phenomenon known as stimulus-specific adaptation (SSA) (168). In this study, it was proposed that, unlike the deviant tone, the repetitive tone activates more inhibitory inputs releasing greater amounts of GABA, which then dampens the firing rate of the SSA neuron via its extrasynaptic GABABRs. While receptor block does not affect the onset of adaptation, suggesting that receptors are not involved in the generation of SSA (168), these observations provide evidence that GABABRs may serve as modulators of predictive coding in the IC.
5 The role of subcortical GABABRs in pathological conditions of the auditory systemDysfunction of the GABABR, an important modulator of cellular excitability, is expected to be part of the mechanisms underlying the neuronal hyperactivity that accompanies some of the known auditory pathologies. Examples of such conditions include noise-induced hearing loss caused by sudden acoustic trauma or prolonged exposure to noise levels above 85 dB (183). It is well documented that this overexposure can lead to death of the IHCs and OHCs, reduction of synaptic ribbons, death of spiral ganglion neurons, or degeneration of the auditory nerve, causing cochlear dysfunction and reduced sensory output (184–186). The peripheral deficit is then thought to trigger mechanisms of homeostatic plasticity in the central auditory pathway to regulate its gain and thereby compensate for the amount of neural activity from the cochlea (187–190). Although the mechanisms underlying neuronal gain modification are not fully understood, one possibility is a reduction in synaptic inhibition mediated by neurotransmitters such as GABA and glycine (191). Consistent with this assumption, studies in animal models of acoustic trauma indicated an imbalance between excitation and inhibition in the auditory system due to impaired GABAergic neurotransmission (192–196), including reduced GABABR expression in the auditory brainstem (121, 122). The latter showed that neurons in the molecular and fusiform layers of the DCN exhibit reduced GABABR density in mice after acoustic trauma, presumably as a result of receptor internalization due to its phosphorylation by protein kinase C gamma (121, 122). This is consistent with the increased excitability of these neurons in animals with noise-induced hearing loss (90, 197, 198). Thus, changes in GABAB function may be part of the mechanisms underlying maladaptive plasticity in the DCN, which is known to lead to hyperactivity of DCN neurons and the development of tinnitus (199, 200). Changes in receptor expression or distribution could also occur in other regions of the subcortical auditory pathway, as suggested by trauma-induced changes in temporal and binaural processing or adaptive coding of sound stimuli, processes that require proper GABABR function (165, 193, 201–204).
Tinnitus, phantom perception in the absence of sound stimuli, is another example of audiological conditions involving hyperactivity of neurons in the auditory pathway (see (90, 198, 205, 206) for reviews). It is generally ignited by hearing loss and very often by noise exposure (207) and, accordingly, animal models of tinnitus show reduced GABA/glycinergic inhibition in various regions of the auditory system (reviewed in (206, 208). Initial work focusing on the relationship between GABABRs and tinnitus explored the possibility of compensating for the loss of GABAergic inhibition by pharmacological activation of receptors with baclofen (165, 209). The results show that baclofen is a potent modulator of neuronal excitability in the ascending auditory pathway that dose-dependently reduces behavioral symptoms of chronic tinnitus in an animal model of acoustic trauma. Later work then indicated a closer link between the receptor and tinnitus by finding that intraperitoneal injection of sodium salicylate, which is known to elicit behavioral measures of tinnitus in animal studies (210), reduced levels of GABAB1 and GABAB2 subunits in the rat IC (211). These observations suggested that baclofen could represent a potentially effective agent in the treatment of tinnitus. However, this assumption failed to be demonstrated in a clinical study of this substance in patients with subjective tinnitus, in which no significant difference was found between the drug and placebo groups (212). Smith et al. (213) addressed the reasons for this failure and suggested that the efficacy of novel GABABR agonists, which do not have the undesirable side effects of baclofen, should be investigated against tinnitus. It is also likely that tinnitus-associated changes in GABABR expression occur only in selected cell subpopulations, as has been proposed, for example, for the inhibitory interneurons of the DCN, cartwheel cells (121). Therefore, more specific substances that selectively modulate the activity of GABABRs in specific cell types or compartments should be found for effective treatment. Interestingly, a variation in KCTD12 gene has been identified as a risk modifier in chronic tinnitus (214). As mentioned above, KCTD12 proteins exhibit multiple modulatory effects on GABABR activity and are expressed in both the peripheral and central parts of the auditory system (11, 59). Their high expression in the stria vascularis suggests their possible involvement in the processes responsible for cochlear K+ transport necessary for the maintenance of the endocochlear potential (11, 85). Therefore, changes in KCTD12 action could contribute to sensorineural hearing loss due to impaired cochlear K+ homeostasis (215, 216) and impaired GABAB function in the auditory pathway, leading to tinnitus. KCTD12 may thus represent a potential target in the therapy of hearing disorders, and it is therefore essential to understand both the mechanisms of its effects in the auditory system and its role in animal models of tinnitus.
6 Concluding remarksThe above data illustrate the involvement of GABABRs in mechanisms that are important for proper hearing function in mammals. These include both peripheral and central processes such as cochlear amplification, sound source localization, temporal processing, adaptive coding, or suppression of spontaneous activity. Because of their relatively slow kinetics, GABABRs are thought to primarily regulate the overall excitability of neurons and thus determine the gain in signal processing. The relatively broad involvement of receptors in auditory processing could then be due to their specific localization in neuronal circuits and the multitude of interacting proteins that serve as their effectors or through which other signaling pathways modulate GABABR activity and thus neuronal responses. The presence of numerous modulation sites on the subunits of GABABRs and their effectors also provides an opportunity to influence receptor responses with pharmacological agents, for example those that restore GABAB function in certain pathological conditions of the auditory system.
As discussed in Section 5, noise or salicylate-induced hearing loss may be accompanied by changes in central gain, and it is likely that attenuation of GABABR signaling contributes to this. Similarly, GABABRs seem to be involved in mechanisms of age-related hearing loss, as suggested by the decline in their levels in the auditory pathway of aging rats (163, 217). By increasing the severity of the auditory deficit, reduced GABABR expression could also indirectly promote pathophysiological processes in other, non-auditory parts of the brain. This assumption is based on the fact that the auditory pathway contains numerous inputs from other sensory systems (see also Sections 4.1 and 4. 3), through which it receives neuromodulation that affects sound processing, and through which it in turn elicits auditory-driven neural responses in non-auditory areas (see (161, 218, 219) for reviews and (220) for recent evidence). Growing evidence suggests a strong association between hearing impairment and cognitive decline, and age-related hearing loss is considered one of the greatest risk factors for the development of dementias, including Alzheimer’s disease (221, 222). The mechanisms linking these states are not entirely clear, but they appear to involve a cascade of events altering the activity of neural circuits in areas important for cognitive function, such as the hippocampus (223–226). Therefore, changes in the activity of auditory neurons due to GABABR dysfunction may potentially play a role in these processes. Hearing loss could be widely treated with hearing aids or cochlear implants and is therefore considered a modifiable risk factor (225), but treatment targeting GABABRs in the auditory pathway could represent a pharmacological alternative. However, the mere systemic use of stimulators of GABABR or its effectors could lead to contradictory effects, as pharmacological inhibition of GABABRs or their G-protein-dependent signaling has been shown to improve hippocampus-dependent memory and decision making, respectively (227–229). This suggests the need for new specific agents targeting GABABRs expressed in the auditory system. The current detailed knowledge of GABABR structure supports advances in pharmacological methods, enabling the investigation of receptor function, as well as the development of new drugs, such as specific allosteric modulators and peptide-based inhibitors that target protein-protein interactions in the receptor complex (230, 231).
Author contributionsRT wrote the paper with input from the other authors. All authors contributed to the article and approved the submitted version.
FundingThis study was supported by grants from the Grant Agency of the Czech Republic (21-17085S), the Charles University Grant Agency (42119), and by the project LX22NPO5107 (MEYS): Financed by EU—Next Generation EU.
AcknowledgmentsWe thank Jan Setnička for technical assistance.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References4. Kulik Á., Booker SA, Vida I. Differential distribution and function of GABA(B)Rs in somato-dendritic and axonal compartments of principal cells and interneurons in cortical circuits. Neuropharmacology (2018) 136:80–91. doi: 10.1016/j.neuropharm.2017.10.018
PubMed Abstract | CrossRef Full Text | Google Scholar
8. Benke D. GABAB receptor trafficking and interacting proteins: targets for the development of highly specific therapeutic strategies to treat neurological disorders? Biochem Pharmacol (2013) 86:1525–30. doi: 10.1016/j.bcp.2013.09.016
PubMed Abstract | CrossRef Full Text | Google Scholar
9. Filip M, Frankowska M, Sadakierska-Chudy A, Suder A, Szumiec L, Mierzejewski P, et al. GABAB receptors as a therapeutic strategy in substance use disorders: focus on positive allosteric modulators. Neuropharmacology (2015) 88:36–47. doi: 10.1016/j.neuropharm.2014.06.016
PubMed Abstract | CrossRef Full Text | Google Scholar
10. Jacobson LH, Vlachou S, Slattery DA, Li X, Cryan JF. The gamma-aminobutyric acid B receptor in depression and reward. Biol Psychiatry (2018) 83:963–76. doi: 10.1016/j.biopsych.2018.02.006
PubMed Abstract | CrossRef Full Text | Google Scholar
11. Resendes BL, Kuo SF, Robertson NG, Giersch ABS, Honrubia D, Ohara O, et al. Isolation from cochlea of a novel human intronless gene with predominant fetal expression. J Assoc Res Otolaryngol JARO (2004) 5:185–202. doi: 10.1007/s10162-003-4042-x
CrossRef Full Text | Google Scholar
12. Juiz JM, Albin RL, Helfert RH, Altschuler RA. Distribution of GABAA and GABAB binding sites in the cochlear nucleus of the Guinea pig. Brain Res (1994) 639:193–201. doi: 10.1016/0006-8993(94)91730-2
PubMed Abstract | CrossRef Full Text | Google Scholar
13. Fubara BM, Casseday JH, Covey E, Schwartz-Bloom RD. Distribution of GABAA, GABAB, and glycine receptors in the central auditory system of the big brown bat, Eptesicus fuscus. J Comp Neurol (1996) 369:83–92. doi: 10.1002/(SICI)1096-9861(19960520)369:1<83::AID-CNE6>3.0.CO;2-G
PubMed Abstract | CrossRef Full Text | Google Scholar
14. Charles KJ, Evans ML, Robbins MJ, Calver AR, Leslie RA, Pangalos MN. Comparative immunohistochemical localisation of GABA(B1a), GABA(B1b) and GABA(B2) subunits in rat brain, spinal cord and dorsal root ganglion. Neuroscience (2001) 106:447–67. doi: 10.1016/S0306-4522(01)00296-2
PubMed Abstract | CrossRef Full Text | Google Scholar
15. Luján R, Shigemoto R, Kulik A, Juiz JM. Localization of the GABAB receptor 1a/b subunit relative to glutamatergic synapses in the dorsal cochlear nucleus of the rat. J Comp Neurol (2004) 475:36–46. doi: 10.1002/cne.20160
PubMed Abstract | CrossRef Full Text | Google Scholar
16. Jamal L, Zhang H, Finlayson PG, Porter LA, Zhang H. The level and distribution of the GABA(B)R2 receptor subunit in the rat's central auditory system. Neuroscience (2011) 6:243–56. doi: 10.1016/j.neuroscience.2011.02.050
留言 (0)