
記住我
Vagus nerve stimulation has emerged as a potential treatment modality in the management of migraine (Straube et al., 2015; Grazzi et al., 2016; Tao et al., 2018; Zhang et al., 2021). Previous studies have demonstrated a significant response to acute transcutaneous vagus nerve stimulation (taVNS) in regions of the vagus nerve pathway, including the nucleus tractus solitarius (NTS), raphe nucleus (RN) and locus coeruleus (LC) in both healthy human participants and patients with migraine (Frangos et al., 2015; Yakunina et al., 2017, 2018; Sclocco et al., 2019, 2020; Zhang et al., 2019; Borgmann et al., 2021). However, the modulation effect of repeated taVNS on these key brain stem regions has yet to be examined.
Approximately 80% of vagus nerve (VN) fibers are sensory fibers that relay both somatic and general visceral signals (Yuan and Silberstein, 2016). The afferent vagal nerves primarily project to the nucleus tractus solitarius, which in turn transmits the signal to the other brain stem nodes such as the locus coeruleus, raphe nuclei, and other subcortical and cortical regions (Dorr and Debonnel, 2006; Manta et al., 2009, 2012; Sacca et al., 2022).
Literature suggests that the locus coeruleus and raphe nuclei are also associated with two neurotransmitter systems potentially important in regulating migraine (Saper, 2011; Obata, 2017). The first neurotransmitter system is the noradrenergic system, which plays a major role in arousal, attention, and the stress response (Berridge and Spencer, 2017), as well as long-term synaptic plasticity and pain modulation (Benarroch, 2009a,b). The locus coeruleus has the greatest amount of noradrenaline in the central nervous system and provides extremely widespread noradrenergic innervations of the entire cortex, diencephalon, and many other brain structures (Benarroch, 2009a,b). Noradrenergic neurons in the LC and its terminals in the dorsal reticular nucleus (DRt), medial prefrontal cortex (mPFC), spinal dorsal horn, and trigeminal spinal nucleus caudalis (spVc) have been shown to participate in the development and maintenance of allodynia and hyperalgesia after nerve injury (Taylor and Westlund, 2017), in addition to their well-described role in inhibiting spinal nociceptive transmissions. Thus, the LC may be a critical modulator for both pain inhibition and facilitation.
The second neurotransmitter system involved in regulating the pain experience is the serotonergic neuromodulatory system. Literature suggests that serotonin plays a major role in modulating pain perception. For instance, combined serotonin-norepinephrine reuptake inhibitors (SNRI) are increasingly considered to be the most effective treatment in the patients comorbid with depression and migraine (Goadsby et al., 2017; Burch, 2019). In the central nervous system, serotonergic innervation of the cerebral cortex, subcortical structures, and cerebellum mainly originate from the dorsal and median raphe nuclei (RN) (Ren et al., 2018).
The aim of this study is to explore the modulatory effect of repeated taVNS on the resting-state functional connectivity (rsFC) of the NTS, LC, and RN in migraine patients. We hypothesize that these brain stem targets of vagal afferents may be modulated by repeated taVNS.
Methods Participants70 patients with migraine were recruited and randomized in an equal ratio to receive real or sham taVNS treatments. Migraine was diagnosed by licensed neurobiologists based on the International Classification of Headache Disorders, 2nd Edition (ICHD-II). The study’s protocol was approved by the Institutional Review Board of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine.
The dataset has been previously used to investigate how taVNS stimulation can modulate functional connectivity of subregions of the thalamus (Zhang et al., 2021). This study focuses on how taVNS can modulate the functional connectivity of key regions in the brain stem vagus nerve pathway (NTS, LC, and RN), which has not been published before. Please see supplementary materials or our previous publication (Zhang et al., 2021) for inclusion and exclusion material.
Experimental procedureAll patients underwent two MRI scans (before and after repeated taVNS). The entire study lasted for about 8 weeks, consisting of a baseline period of 4 weeks collecting migraine attack characteristics prior to treatment and the 4-week treatment period. At enrollment, patients were requested to maintain diary records of their headaches for the duration of the study. Each recorded headache entry included onset time, duration, pain intensity (using Visual Analog Scale (VAS) score), accompanying symptoms, and the use of rescue medication if any. Both real and sham taVNS treatments consisted of 12 sessions in a schedule of 3 sessions per week for 4 weeks, with each session lasting for 30 min. A 1 Hz continuous electrical stimulation was applied at the left cymba concha (Badran et al., 2018) for real taVNS treatment, while the sham stimulation site was located on the left tail of the helix (Badran et al., 2018). See supplementary materials for details in experimental procedures and data acquisition.
Functional analysisFunctional data preprocessing and statistical analysis were performed in the SPM12-based toolbox CONN19.c (Whitfield-Gabrieli and Nieto-Castanon, 2012), SUIT (Diedrichsen, 2006), and AFNI software version 17.2.05 (Cox et al., 2017). See supplementary materials for data preprocessing procedures.
The bilateral NTS were defined based on a publicly available template provided by the team of Nikos Priovoulos (Priovoulos et al., 2019). The bilateral RN (combination of median and dorsal raphe) and LC seeds were defined based on the Automatic Anatomical Labeling 3 atlas 3 (AAL) template (Pollak Dorocic et al., 2014). In the first-level analysis, we produced a correlation map for each patient by extracting the BOLD time series from each brainstem seed and computing Pearson’s correlation coefficients between the time series in every seed and all other voxels of the whole brain, respectively. Correlation coefficients were Fisher transformed into ‘Z’ scores to increase normality.
Seed-to-voxel second-level analyses were performed using a mixed-designed ANOVA with treatment (real taVNS vs. sham taVNS) entered as the between-subject factor, time (pre-treatment vs. post-treatment) as the within-subject factor, and age and gender as covariates. We also performed a one-sample t-test to assess the baseline functional connectivity of each seed in all migraine patients. A threshold of voxel-wise p < 0.005 and cluster-level p < 0.05 FDR corrected was applied for second-level analyses.
Based on previous imaging studies, our a prior regions of interest included: (1) downstream targets of vagal afferents in the brain stem: LC and RN (Dorr and Debonnel, 2006; Manta et al., 2009, 2012) and (2) the pain perception and modulation network in migraine: hypothalamus, thalamus, hippocampus, amygdala, insula, anterior cingulate cortex (ACC), medial prefrontal cortex (mPFC) and postcentral gyrus (PoCG) (Frangos et al., 2015; Hachem et al., 2018; Kaniusas et al., 2019; Lee et al., 2019; Silberstein et al., 2020; Ashina et al., 2021). An initial threshold of voxel-wise p < 0.005 was used in all data analysis. To correct for multiple comparisons, Monte Carlo simulations using the 3dFWHMx and 3dClustSim implemented in AFNI were applied for these ROIs (i.e., brain regions listed above, where for each region, the minimum voxel size required for p < 0.05 cluster level p-value correction will be indicated as a k value in the results presented below). For the rest of the brain, a voxel-wise threshold of p < 0.005 with p < 0.05 cluster level FDR correction was applied. A similar analysis was performed in our previous study (Gollub et al., 2018).
To investigate the association between the rsFC change and the primary clinical outcome (number of migraine attack days), we created a sphere with a 2 mm radius using the peak coordinates of the brain regions that produced significant rsFC changes after taVNS and extracted the average z values. Then, we performed a correlation analysis between the rsFC z value changes and the corresponding clinical outcomes across all participants.
Results Clinical outcome59 patients (33 in the taVNS group, 26 in the sham taVNS group) completed two MRI scans before and after 4 weeks of treatment. No patients were excluded due to fMRI head motion correction based on the calculated mean frame displacement (FD) standard.
Compared to the sham taVNS group, patients in the real taVNS treatment group had a significant reduction in the number of migraine attack days ([F (1,57) =5.41, p = 0.024]), headache pain intensity ([F (1,57) = 7.52, p = 0.008]), and migraine attack times (numbers) ([F (1,57) = 6.29, p = 0.015]). There was no significant improvement in MSQ, SAS and SDS evaluation after real taVNS treatment. Detailed patients’ baseline characteristics and clinical outcome measurements were shown in Supplementary Table e1.
Functional connectivity results NTS functional connectivity resultsAmong all migraine patients, baseline functional connectivity of the NTS showed positive connectivity with the bilateral angular gyrus (AG), superior/middle frontal gyrus, posterior cingulate gyrus, fusiform gyrus; left hippocampus/parahippocampus, thalamus, cerebellum, and right lingual gyrus; and negative connectivity with the bilateral dorsal lateral prefrontal cortex (dlPFC), putamen, middle/superior temporal gyrus, supramarginal gyrus, supplementary motor cortex, superior frontal gyrus (SFG), left inferior occipital gyrus (IOG), postcentral gyrus (PoCG), and right precentral gyrus (PreCG). Baseline functional connectivity of the NTS is shown in Supplementary Table e2.
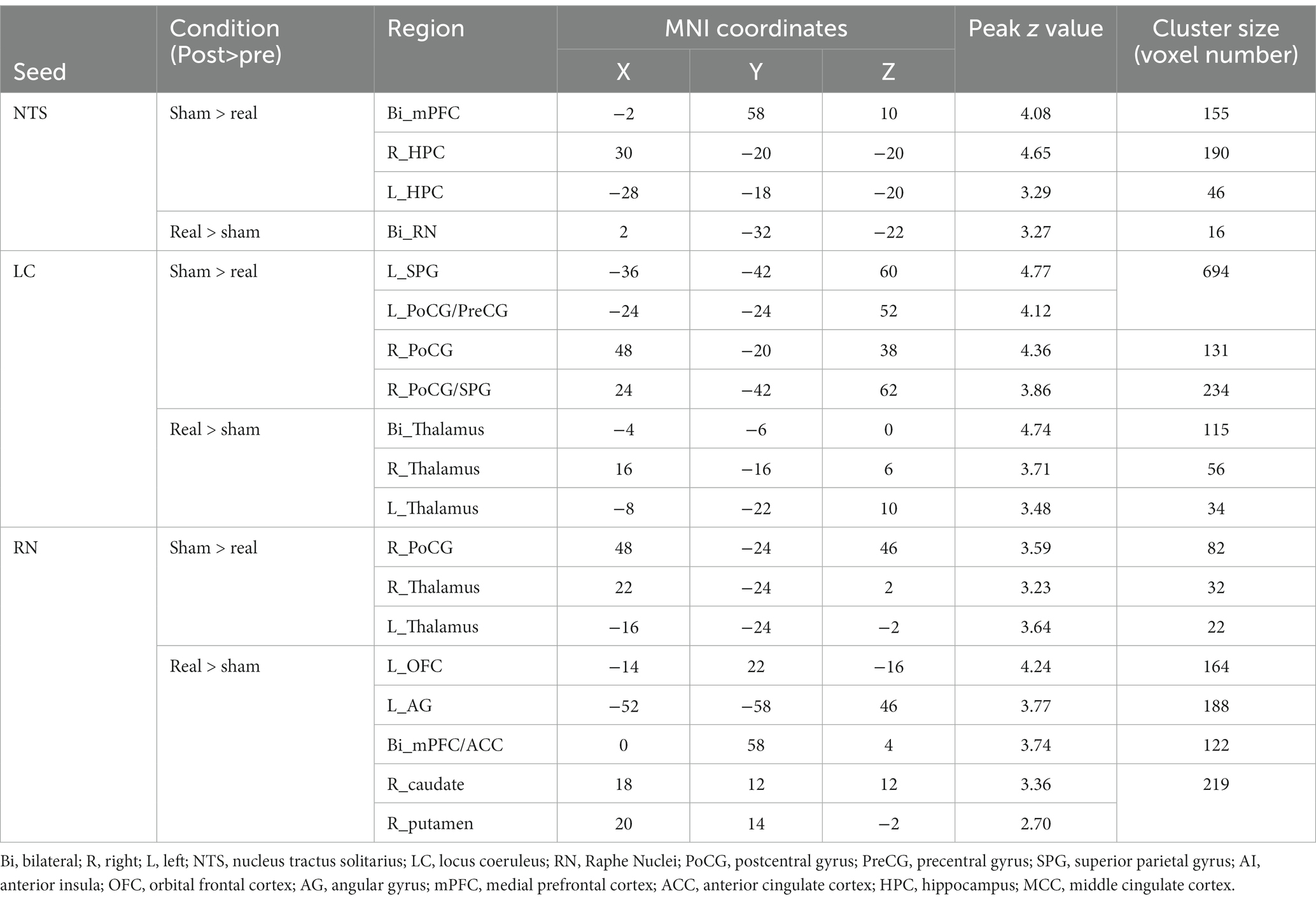
Compared to the sham group, the real taVNS group showed decreased NTS functional connectivity with the bilateral medial prefrontal cortex (mPFC) and bilateral hippocampus (HPC); and increased functional connectivity with the bilateral raphe nuclei after treatment (Table 1 and Figure 1).
Table 1. Seed-based analysis results.
Figure 1. Group comparison results. Bi, bilateral; R, right; L, left; PoCG, postcentral gyrus; PreCG, precentral gyrus; SPG, superior parietal gyrus; AI, anterior insula; OFC, orbital frontal cortex; AG, angular gyrus; mPFC, medial prefrontal cortex; HPC, hippocampus; MCC, middle cingulate cortex.
LC functional connectivity resultsBaseline functional connectivity of the LC showed positive connectivity with the bilateral cerebellum, lingual gyrus, and hippocampus/parahippocampus, and negative connectivity with right occipital fusiform gyrus (Supplementary Table e2).
Compared to the sham group, the real taVNS group showed decreased LC functional connectivity with the bilateral postcentral gyrus (PoCG), superior parietal gyrus (SPG), and left precentral gyrus (PreCG); and increased functional connectivity with the bilateral thalamus after treatment (Table 1 and Figure 1).
RN functional connectivity resultsBaseline functional connectivity of the RN showed positive connectivity with the left thalamus and negative connectivity with the left caudate and right occipital pole (Supplementary Table e2).
Compared to the sham group, the real taVNS group showed decreased RN functional connectivity with the bilateral thalamus and right PoCG; and increased functional connectivity with the bilateral mPFC/anterior cingulate cortex (ACC), left occipital frontal cortex (OFC), left angular gyrus (AG), right caudate, and putamen after treatment (Table 1 and Figure 1).
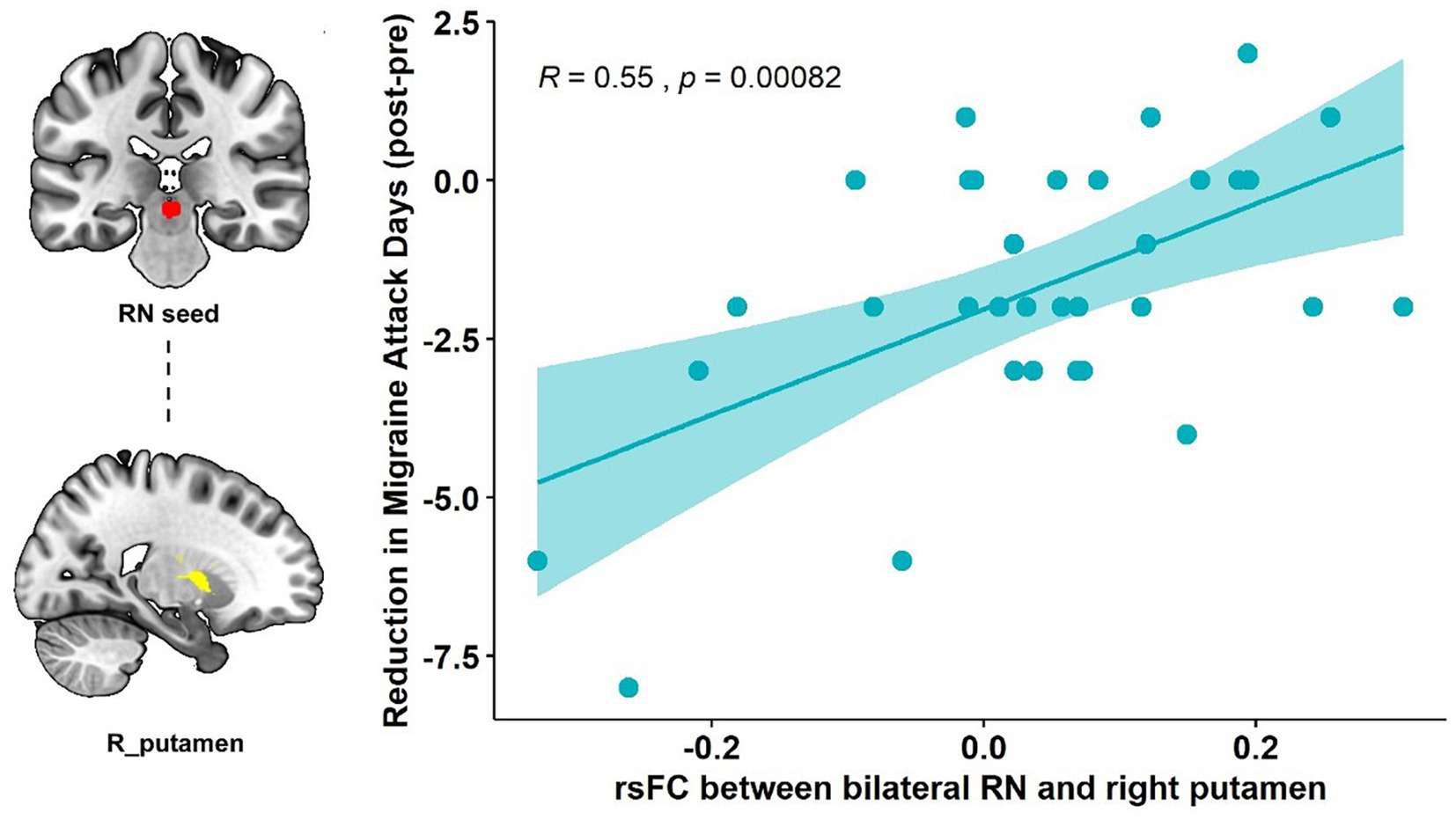
Association with primary clinical outcomePearson’s correlation showed that the RN – right putamen rsFC change was significantly correlated with the primary outcome (reduction of the number of migraine days, R = 0.55, p < 0.001, p = 0.015 after Bonferroni correction) in the real taVNS group after treatment (Figure 2).
Figure 2. Correlation results of the RN-putamen rsFC change and clinical outcomes. The scatterplot indicates the significant correlation between the change in number of migraine attack days and the rsFC change of the bilateral RN and right putamen after real taVNS treatment (R = 0.55, p < 0.001, significant after Bonferroni correction).
There is no significant association between NTS/LC rsFC change and the corresponding primary clinical outcome changes.
DiscussionIn the present study, we investigated the modulatory effects of repeated taVNS on functional connectivity of the NTS, LC and RN in patients with migraine. Comparisons of clinical measurements indicated repeated taVNS could significantly relieve patients’ symptoms. Functional analysis showed that repeated taVNS could modulate the connection between the NTS and key brain regions in the limbic system (bilateral hippocampus), and the functional connectivity of both the LC and RN with brain regions closely relating to pain processing and modulation (bilateral postcentral gyrus/bilateral thalamus/mPFC). Our findings also demonstrated that repeated taVNS could strengthen the functional connectivity between the RN and caudate/putamen, the main dopaminergic projecting targets. The RN-caudate/putamen rsFC change was significantly associated with the reduction in the number of migraine attack days. These findings suggests that repeated taVNS could significantly modulate the vagus nerve pathway in patients with migraine.
As the major target of vagal afferents, the NTS receives direct inputs from the vagus nerve and subsequently projects to other brainstem regions including the LC, RN and parabrachial nucleus, along with the forebrain limbic structures such as the periventricular nucleus of the hypothalamus, the thalamus, the central nucleus of the amygdala, the hippocampus, the bed nucleus of the stria terminalis, and the nucleus accumbens (Zoccal et al., 2014). In the current study, we observed a significantly decreased NTS rsFC with the bilateral mPFC and hippocampus and increased rsFC with the RN after the real taVNS treatments compared to the sham.
The hippocampus is a main structure of the limbic system. Accumulating evidence suggests that limbic system, and the hippocampus alone, is involved in pain processing, pain-related attention, anxiety, and the stress response (Liu and Chen, 2009; Mokhtari et al., 2019; Yu et al., 2020). Both structural and functional abnormalities of the hippocampus have been reported in migraine patients (Biggio et al., 2009). Literature suggests that headache frequency, cumulative number of migraine attacks, anxiety scores, depression scores, and genetic variants are related to hippocampal morphology and functional changes in individuals with migraine (Liu et al., 2018). A previous neuroimaging study found that migraineurs had greater pain-induced activation of the hippocampus, and the activation strength had a significant correlation with headache frequency in migraine patients (Schwedt et al., 2014). Our results suggest that taVNS could modulate the functional connection between the hippocampus and the NTS. This finding is also consistent with a previous study in which the authors found that plasticity within the hippocampus is an important contributor to the vagus nerve response (Biggio et al., 2009).
We found that taVNS could modulate the rsFC of both the LC and RN with brain regions closely relating to pain processing and modulation (PoCG/thalamus/mPFC). In particular, both the LC and RN is associated with a reduced functional connectivity with the PoCG after repeated taVNS treatment. Previous studies have identified involvement of the PoCG in chronic pain (Kong et al., 2013; Ossipov et al., 2014). Particularly, the majority of patients with migraine show somatosensory hypersensitivity (i.e., reduced cutaneous pain thresholds) and other symptoms of somatosensory hypersensitivity (i.e., cutaneous allodynia) during migraine attacks (Schwedt, 2013). The LC sends strong projections to the sensory cortex and has long been posited to play an important role in modulating sensory encoding (Bruinstroop et al., 2012; Vazey et al., 2018).
As the major source of serotonergic projections, the raphe nucleus plays a role in the regulation of multiple functional systems, including the somatosensory system (Hornung, 2003). Our results suggest that repeated taVNS may regulate the perception of nociceptive inputs in patients with migraine by interfering with the connection between LC/RN and PoCG. These connectivity changes may alter the projection of noradrenergic and serotonergic neurotransmitters in the brain stem to the somatosensory cortex.
The thalamus is a major center in the brain for processing nociceptive information and relays this information to cortical regions for further processing (Lenz et al., 2004). Evidence of activation of the thalamus during migraines is clear and well established (Bahra et al., 2001; Afridi et al., 2005; Tu et al., 2019). It is worth noting that our results showed that repeated taVNS could modulate the rsFC of both the LC and RN with the thalamus in patients with migraine. A previous study found that both noradrenergic and serotonergic fibers innervate the thalamus, and the origins of these projections are primarily located at the LC and RN (Westlund et al., 1991). We also found that taVNS can significantly modulate the connectivity between the thalamus and sensory-motor cortex (Zhang et al., 2021). Interestingly, we found that after repeated taVNS, the rsFC of the thalamus was increased with the LC but decreased with the RN. As the noradrenergic and serotonergic fibers project to multiple nuclei of the thalamus, involving both excitatory and inhibitory effects (Varela, 2014), future studies are needed to investigate more details on how taVNS may affect the LC and RN projections to the thalamus.
We also found that taVNS could significantly modulate the rsFC between the RN and bilateral mPFC / ACC in patients with migraine. The mPFC is a crucial brain region that integrates information from multiple cortical and subcortical areas and converges updated outputs to downstream structures. In addition, the mPFC / ACC is also a key region of the descending pain modulation system (Kong et al., 2010). The functional and structural alterations of the mPFC in patients with migraine have been reported (Jin et al., 2013; Soheili-Nezhad et al., 2019). It was considered to play an essential role in regulating pain-related emotion (depression and anxiety) and cognition in patients with migraine (Ma et al., 2018; Tu et al., 2019; Zhang et al., 2019). A previous study showed that migraine is associated with increased PAG – mPFC connectivity, and the increased connectivity decreased after acupuncture treatments (Li et al., 2016). We also found taVNS can modulate the PAG – ACC/mPFC connectivity in patient with migraine (Cao et al., 2021). Interestingly, we explored the association between the RN-mPFC rsFC change and changes in depression scores in the repeated taVNS treatment group. The results showed that these two factors are highly correlated, with a p value of 0.0038 (R = −0.49). Our result suggests that taVNS may hold the potential to relieve both headache and depression in patients comorbid with migraine and depression. This is consistent with our previous studies showing that taVNS can also reduce symptoms in patients with depression (Kong et al., 2018; Li et al., 2022).
Our results showed that repeated taVNS significantly altered the functional connectivity between the RN and basal ganglia subregions, specifically the right putamen and caudate. This connectivity change is associated with a reduction in the number of migraine attack days, i.e., the higher the functional strength between RN and right putamen, the greater the reduction in the number of attack days in migraine patients.
Literature has suggested that the basal ganglia may be involved in the sensory, emotional / cognitive, and endogenous/modulatory domains of pain processing (Borsook et al., 2010). In particular, the putamen is one of the major sites of cortical input into basal ganglia loops and is frequently activated during pain (Starr et al., 2011). Dysfunctions of the monoamine systems, especially the dopaminergic and serotonergic projections to the basal ganglia, were reported to correlate with individual ratings of the sensory and unpleasant qualities of the experience of pain(Scott et al., 2006) and might be the early result of chronic maladaptation to persistent pain (Sagheddu et al., 2015). Thus, we speculate that the repeated administration of taVNS might modulate pain perception attributed to the serotonin activation in the putamen projected from the RN in patients with migraine.
There are several limitations to this study. Firstly, the current study was conducted over a 4-week period and therefore the effects observed can only represent short-and mid-term effects; a study with a longer duration is needed to investigate long-term effects. Secondly, the sham stimulation was applied at the tail of the helix with the same electrical stimulation parameters, which might also activate the sensory related cortex. Caution needs to be taken in the interpretation of the comparison results of the rsFC change between the two groups, especially when it comes to the upregulation of somatosensory cortex. Future studies are needed to validate / replicate our findings.
In conclusion, we found that repeated taVNS can modulate the connection between the key nodes of the brain stem vagus nerve pathway with the limbic system and pain perception and modulation networks. These findings suggest that repeated taVNS may yield pain relief through modulating of the NTS, LC, and RN connections with the above regions, which may further affect their synaptic plasticity and neurotransmitter release associated with migraine.
Data availability statementThe raw data supporting the conclusions of this article will be made available per request.
Ethics statementThe studies involving human participants were reviewed and approved by the Institutional Review Board of the Second Affiliated Hospital of Guangzhou University of Chinese Medicine. The patients/participants provided their written informed consent to participate in this study.
Author contributionsYZ, BL, and JK: design and conceptualized study. YH and JK: analyze and interpret the data. YZ, HL, ZY, XL, XH, and WC: acquisition of data. YH, SH, TC-Z, and JK: manuscript preparation. YZ and BL: funding acquisition. All authors contributed to the article and approved the submitted version.
FundingThis study was supported by the Medical Scientific Research Foundation of Guangdong Province of China (A2017234) and the Administration of Traditional Chinese Medicine of Guangdong Province of China (20182047).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnmol.2023.1160006/full#supplementary-material
Footnotes ReferencesAfridi, S. K., Giffin, N. J., Kaube, H., Friston, K. J., Ward, N. S., Frackowiak, R. S. J., et al. (2005). A positron emission tomographic study in spontaneous migraine. Arch. Neurol. 62, 1270–1275. doi: 10.1001/archneur.62.8.1270
PubMed Abstract | CrossRef Full Text | Google Scholar
Ashina, S., Bentivegna, E., Martelletti, P., and Eikermann-Haerter, K. (2021). Structural and functional brain changes in migraine. Pain Ther. 10, 211–223. doi: 10.1007/s40122-021-00240-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Badran, B. W., Brown, J. C., Dowdle, L. T., Mithoefer, O. J., LaBate, N. T., Coatsworth, J., et al. (2018). Tragus or cymba conchae? Investigating the anatomical foundation of transcutaneous auricular vagus nerve stimulation (taVNS). Brain Stimul. 11, 947–948. doi: 10.1016/j.brs.2018.06.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Bahra, A., Matharu, M. S., Buchel, C., Frackowiak, R. S. J., and Goadsby, P. J. (2001). Brainstem activation specific to migraine headache. Lancet 357, 1016–1017. doi: 10.1016/S0140-6736(00)04250-1
CrossRef Full Text | Google Scholar
Benarroch, E. E. (2009a). The locus ceruleus norepinephrine system-functional organization and potential clinical significance. Neurology 73, 1699–1704. doi: 10.1212/WNL.0b013e3181c2937c
PubMed Abstract | CrossRef Full Text | Google Scholar
Benarroch, E. E. (2009b). The locus ceruleus norepinephrine system: functional organization and potential clinical significance. Neurology 73, 1699–1704. doi: 10.1212/WNL.0b013e3181c2937c
CrossRef Full Text | Google Scholar
Berridge, C. W., and Spencer, R. C. (2017). “Chapter 4 - noradrenergic control of arousal and stress” in Stress: neuroendocrinology and neurobiology. ed. G. Fink (San Diego: Academic Press), 39–47.
Biggio, F., Gorini, G., Utzeri, C., Olla, P., Marrosu, F., Mocchetti, I., et al. (2009). Chronic vagus nerve stimulation induces neuronal plasticity in the rat hippocampus. Int. J. Neuropsychopharmacol. 12, 1209–1221. doi: 10.1017/S1461145709000200
PubMed Abstract | CrossRef Full Text | Google Scholar
Borgmann, D., Rigoux, L., Kuzmanovic, B., Edwin Thanarajah, S., Münte, T. F., Fenselau, H., et al. (2021). Technical note: modulation of fMRI brainstem responses by transcutaneous vagus nerve stimulation. NeuroImage 244:118566. doi: 10.1016/j.neuroimage.2021.118566
PubMed Abstract | CrossRef Full Text | Google Scholar
Borsook, D., Upadhyay, J., Chudler, E. H., and Becerra, L. (2010). A key role of the basal ganglia in pain and analgesia--insights gained through human functional imaging. Mol. Pain 6:27. doi: 10.1186/1744-8069-6-27
CrossRef Full Text | Google Scholar
Bruinstroop, E., Cano, G., Vanderhorst, V. G. J. M., Cavalcante, J. C., Wirth, J., Sena-Esteves, M., et al. (2012). Spinal projections of the A5, A6 (locus coeruleus), and A7 noradrenergic cell groups in rats. J. Comp. Neurol. 520, 1985–2001. doi: 10.1002/cne.23024
PubMed Abstract | CrossRef Full Text | Google Scholar
Burch, R. (2019). Antidepressants for preventive treatment of migraine. Curr. Treat. Options Neurol. 21:18. doi: 10.1007/s11940-019-0557-2
CrossRef Full Text | Google Scholar
Cao, J., Zhang, Y., Li, H., Yan, Z., Liu, X., Hou, X., et al. (2021). Different modulation effects of 1 Hz and 20 Hz transcutaneous auricular vagus nerve stimulation on the functional connectivity of the periaqueductal gray in patients with migraine. J. Transl. Med. 19:354. doi: 10.1186/s12967-021-03024-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Cox, R. W., Chen, G., Glen, D. R., Reynolds, R. C., and Taylor, P. A. (2017). fMRI clustering and false-positive rates. Proc. Natl. Acad. Sci. U. S. A. 114, E3370–e3371. doi: 10.1073/pnas.1614961114
PubMed Abstract | CrossRef Full Text | Google Scholar
Diedrichsen, J. (2006). A spatially unbiased atlas template of the human cerebellum. NeuroImage 33, 127–138. doi: 10.1016/j.neuroimage.2006.05.056
CrossRef Full Text | Google Scholar
Dorr, A. E., and Debonnel, G. (2006). Effect of vagus nerve stimulation on serotonergic and noradrenergic transmission. J. Pharmacol. Exp. Ther. 318, 890–898. doi: 10.1124/jpet.106.104166
PubMed Abstract | CrossRef Full Text | Google Scholar
Frangos, E., Ellrich, J., and Komisaruk, B. R. (2015). Non-invasive access to the Vagus nerve central projections via electrical stimulation of the external ear: fMRI evidence in humans. Brain Stimul. 8, 624–636. doi: 10.1016/j.brs.2014.11.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Goadsby, P. J., Holland, P. R., Martins-Oliveira, M., Hoffmann, J., Schankin, C., and Akerman, S. (2017). Pathophysiology of migraine: a disorder of sensory processing. Physiol. Rev. 97, 553–622. doi: 10.1152/physrev.00034.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Gollub, R. L., Kirsch, I., Maleki, N., Wasan, A. D., Edwards, R. R., Tu, Y., et al. (2018). A functional neuroimaging study of expectancy effects on pain response in patients with knee osteoarthritis. J. Pain 19, 515–527. doi: 10.1016/j.jpain.2017.12.260
PubMed Abstract | CrossRef Full Text | Google Scholar
Grazzi, L., Egeo, G., Calhoun, A. H., McClure, C. K., Liebler, E., and Barbanti, P. (2016). Non-invasive Vagus nerve stimulation (nVNS) as mini-prophylaxis for menstrual/menstrually related migraine: an open-label study. J. Headache Pain 17:91. doi: 10.1186/s10194-016-0684-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Hachem, L. D., Wong, S. M., and Ibrahim, G. M. (2018). The vagus afferent network: emerging role in translational connectomics. Neurosurg. Focus. 45:E2. doi: 10.3171/2018.6.FOCUS18216
CrossRef Full Text | Google Scholar
Hornung, J. P. (2003). The human raphe nuclei and the serotonergic system. J. Chem. Neuroanat. 26, 331–343. doi: 10.1016/j.jchemneu.2003.10.002
CrossRef Full Text | Google Scholar
Jin, C., Yuan, K., Zhao, L., Zhao, L., Yu, D., von Deneen, K. M., et al. (2013). Structural and functional abnormalities in migraine patients without aura. NMR Biomed. 26, 58–64. doi: 10.1002/nbm.2819
CrossRef Full Text | Google Scholar
Kaniusas, E., Kampusch, S., Tittgemeyer, M., Panetsos, F., Gines, R. F., Papa, M., et al. (2019). Current directions in the auricular Vagus nerve stimulation I – a physiological perspective. Front. Neurosci. 13:854. doi: 10.3389/fnins.2019.00854
CrossRef Full Text | Google Scholar
Kong, J., Fang, J., Park, J., Li, S., and Rong, P. (2018). Treating depression with transcutaneous auricular Vagus nerve stimulation: state of the art and future perspectives. Front. Psych. 9:20. doi: 10.3389/fpsyt.2018.00020
PubMed Abstract | CrossRef Full Text | Google Scholar
Kong, J., Spaeth, B., Wey, H. Y., Cheetham, A., Cook, A. H., Jensen, K., et al. (2013). S1 is associated with chronic low back pain: a functional and structural MRI study. Mol. Pain 9:43. doi: 10.1186/1744-8069-9-43
CrossRef Full Text | Google Scholar
Kong, J., Tu, P. C., Zyloney, C., and Su, T. P. (2010). Intrinsic functional connectivity of the periaqueductal gray, a resting fMRI study. Behav. Brain Res. 211, 215–219. doi: 10.1016/j.bbr.2010.03.042
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, M. J., Park, B. Y., Cho, S., Kim, S. T., Park, H., and Chung, C. S. (2019). Increased connectivity of pain matrix in chronic migraine: a resting-state functional MRI study. J. Headache Pain 20:29. doi: 10.1186/s10194-019-0986-z
PubMed Abstract | CrossRef Full Text | Google Scholar
Lenz, F. A., Weiss, N., Ohara, S., Lawson, C., and Greenspan, J. D. (2004). “Chapter 6 the role of the thalamus in pain” in Supplements to clinical neurophysiology. eds. M. Hallett, L. H. Phillips, D. L. Schomer, and J. M. Massey, vol. 57 (Radarweg, Amsterdam: Elsevier), 50–61.
Li, Z., Liu, M., Lan, L., Zeng, F., Makris, N., Liang, Y., et al. (2016). Altered periaqueductal gray resting state functional connectivity in migraine and the modulation effect of treatment. Sci. Rep. 6:20298. doi: 10.1038/srep20298
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, S., Rong, P., Wang, Y., Jin, G., Hou, X., Li, S., et al. (2022). Comparative effectiveness of transcutaneous auricular Vagus nerve stimulation vs citalopram for major depressive disorder: a randomized trial. Neuromodulation 25, 450–460. doi: 10.1016/j.neurom.2021.10.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, M.-G., and Chen, J. (2009). Roles of the hippocampal formation in pain information processing. Neurosci. Bull. 25, 237–266. doi: 10.1007/s12264-009-0905-4
CrossRef Full Text | Google Scholar
Liu, H.-Y., Chou, K.-H., and Chen, W.-T. (2018). Migraine and the Hippocampus. Curr. Pain Headache Rep. 22:13. doi: 10.1007/s11916-018-0668-6
CrossRef Full Text | Google Scholar
Ma, M., Zhang, J., Chen, N., Guo, J., Zhang, Y., and He, L. (2018). Exploration of intrinsic brain activity in migraine with and without comorbid depression. J. Headache Pain 19:48. doi: 10.1186/s10194-018-0876-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Manta, S., Dong, J., Debonnel, G., and Blier, P. (2009). Enhancement of the function of rat serotonin and norepinephrine neurons by sustained vagus nerve stimulation. J. Psychiatry Neurosci. 34, 272–280.
PubMed Abstract | Google Scholar
Manta, S., El Mansari, M., Debonnel, G., and Blier, P. (2012). Electrophysiological and neurochemical effects of long-term vagus nerve stimulation on the rat monoaminergic systems. Int. J. Neuropsychopharmacol. 16, 459–470. doi: 10.1017/S1461145712000387
PubMed Abstract | CrossRef Full Text | Google Scholar
Mokhtari, T., Tu, Y., and Hu, L. (2019). Involvement of the hippocampus in chronic pain and depression. Brain Sci. Adv. 5, 288–298. doi: 10.26599/BSA.2019.9050025
CrossRef Full Text | Google Scholar
Ossipov, M. H., Morimura, K., and Porreca, F. (2014). Descending pain modulation and chronification of pain. Curr. Opin. Support. Palliat. Care 8, 143–151. doi: 10.1097/SPC.0000000000000055
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollak Dorocic, I., Fürth, D., Xuan, Y., Johansson, Y., Pozzi, L., Silberberg, G., et al. (2014). A whole-brain atlas of inputs to serotonergic neurons of the dorsal and median raphe nuclei. Neuron 83, 663–678. doi: 10.1016/j.neuron.2014.07.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Priovoulos, N., Poser, B. A., Ivanov, D., Verhey, F. R. J., and Jacobs, H. I. L. (2019). In vivo imaging of the nucleus of the solitary tract with magnetization transfer at 7 tesla. NeuroImage 201:116071. doi: 10.1016/j.neuroimage.2019.116071
CrossRef Full Text | Google Scholar
Ren, J., Friedmann, D., Xiong, J., Liu, C. D., Ferguson, B. R., Weerakkody, T., et al. (2018). Anatomically defined and functionally distinct dorsal raphe serotonin sub-systems. Cells 175, 472–487.e20. doi: 10.1016/j.cell.2018.07.043
PubMed Abstract | CrossRef Full Text | Google Scholar
Sacca, V., Zhang, Y., Cao, J., Li, H., Yan, Z., Ye, Y., et al. (2022). Evaluation of the modulation effects evoked by different transcutaneous auricular Vagus nerve stimulation frequencies along the central Vagus nerve pathway in migraines: a functional magnetic resonance imaging study. Neuromodulation: journal of the international Neuromodulation. Society 26, 620–628. doi: 10.1016/j.neurom.2022.08.459
留言 (0)