


記住我
Acanthamoeba spp. are ubiquitous free-living amoebae occurring worldwide in water environments and soil, but they can also be isolated from dust and the air. They are facultative pathogens and can cause different diseases, importantly, the so-called Acanthamoeba keratitis (AK), a severe infection of the cornea, most commonly observed in contact lens wearers. The incidence of AK has increased within the past decades, which may be attributed to the increasing number of contact lens users, but also to advances in diagnostics (Chawla et al., 2014; Randag et al., 2019; List et al., 2021). Acanthamoeba spp. may also cause granulomatous amoebic encephalitis, a fatal disease of the central nervous system, and other disseminating infections, mainly in immunocompromised individuals (Morrison et al., 2016). Treating these infections is challenging, as no specific drugs are currently available (Siddiqui et al., 2014; Lorenzo-Morales et al., 2015; Loufouma Mbouaka et al., 2021).
The pathomechanism of these amoebae still remains incompletely understood, despite significant progress made in recent years (Clarke and Niederkorn, 2006; Khan, 2006; Walochnik and Duchêne, 2016). Adhesion to the host cells is mediated by a mannose-binding protein (MBP) and other adhesion proteins, triggering the release of proteases, mainly of the serine and metalloprotease type. The immune reaction of the host is characterized by neutrophil migration and macrophage activation resulting in the release of proinflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6). In AK, secretory IgA antibodies in the tears can prevent binding of the Acanthamoeba trophozoites (Mattana et al., 2002; Garate et al., 2004; Walochnik and Duchêne, 2016). However, only a comparably small percentage of environmental Acanthamoeba isolates are able to lyse human cells and cytopathogenicity is known to decline during long-term axenic culture – but can also be enhanced by mouse passage or serial passage over human cell lines (Mazur and Hadaś, 1994; Koehsler et al., 2009). Cell viability and cytotoxicity assays are useful tools to determine the cytotoxic effects of Acanthamoeba on human cells because they measure in vitro modifications at the cellular and metabolic levels by detecting structural changes such as loss of membrane integrity or physiological and biochemical responses associated with non-viable and viable cells, respectively (Riss et al., 2004). They are typically used in drug discovery screening to assess the effect of a compound on cell proliferation (Riss et al., 2004). However, depending on the research aims and owing to the limitations of these assays, their use may be challenging. Recently, it was demonstrated that some of these assays, when used to study and evaluate host–pathogen interactions, may interfere with the pathogen and lead to an inaccurate estimation of pathogen cytotoxicity and their effects on host cells during and after co-culture (Van den Bossche et al., 2020). Thus, it is crucial to identify or develop and validate reliable methods and models to study such interactions and to avoid any interference with the culture medium or the pathogen in co-culture.
The present study aimed to evaluate and compare the most widely used cell viability assays for their usefulness to assess the cytotoxicity of Acanthamoeba spp. on human corneal epithelial cells (HCECs) during co-culture. Cell morphology, viability and integrity were evaluated by phase contrast microscopy.
2. Materials and methods 2.1. Acanthamoeba strainsThe non-pathogenic environmental isolate strain Acanthamoeba castellanii Neff (ATCC 50373) and two pathogenic isolates from patients with keratitis, strains 1BU and strain SIN20, isolated in 1998 and 2020, respectively, all belonging to the T4 genotype group, were used in this study. The strains were maintained on non-nutrient agar plates coated with Escherichia coli. Prior to the experiments, all strains were sub-cultured and grown axenically at 34°C in peptone, yeast extract, and glucose (PYG) medium containing 10 g proteose peptone, 10 g yeast extract, 5 g NaCl, 5 g glucose, 0.7 g Na2HPO4, and 0.7 g KH2PO4 per liter.
2.2. Human corneal epithelial cells (HCECs)Cells and media components were purchased from Innoprot (Derio, Bizkaia, Spain). Immortalized human corneal epithelial cells (HCECs; P10871-IM) were sub-cultured in corneal epithelial cell medium (CEpiCM, P60189) containing the 5% fetal bovine serum (FBS), 1% epithelial cell growth supplement (ECGS) and 1% penicillin/streptomycin at 37°C and 5% CO2. T75 flasks and 96-well plates were coated with a thin layer of type I collagen (P8188) to enhance cell attachment and proliferation. Prior to the assays, different concentrations of HCECs were assessed to determine the optimal concentration for the subsequent experiments.
2.3. Acanthamoeba–HCEC co-cultureAt least three independent experiments were performed in triplicates. Before the assays, 1 × 104 HCECs per well were seeded in a 96-well plate and incubated overnight. Then, the medium was replaced with a serum-free medium, and cells were maintained at 34°C and 5% CO2 during the co-culture to mimic the conditions of the human eye. Amoebae were added into the wells at different ratios or multiplicity of infection (MOI 1, MOI 2, and MOI 3; see Table 1) and incubated for 2, 4, 6, and 8 h. Wells containing only the HCECs were considered as positive controls, with a percentage of viability close to 100% for the following assays.
Table 1. Multiplicity of infection (MOI) used for cytotoxicity assays.
2.4. Cell viability and cytotoxicity assays 2.4.1. Lactate dehydrogenase assayLactate dehydrogenase (LDH) assay is a colorimetric method used to assess cytotoxicity and quantify cell viability. The damage to the plasma membrane allows the release of LDH from the intracellular environment into the cell culture medium and can be quantified using a coupled enzymatic reaction. The LDH activity was determined using the CyQUANT™ LDH Cytotoxicity Assay Kit (Thermo Fisher Scientific, Eugene, OR, USA), and strictly following the manufacturers’ instructions (Thermo Fisher, 2019). Simultaneously, the LDH positive control was established by adding the same volume of lysis buffer to samples containing only HCECs or Acanthamoeba alone. The plate was incubated at room temperature and protected from light for 30 min. Then, the absorbance was measured at 490 nm using a microplate absorbance spectrophotometer (Anthos Labtec Instruments HT2, Salzburg, Austria). The percentage of cytotoxicity was determined following the formula provided with the LDH assay kit and then converted to relative cell viability.
2.4.2. MTS assayThe MTS assay is a colorimetric method used to assess cell viability. Viable cells reduce the yellow tetrazolium compound [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt; MTS] to soluble formazan, which is purple. CellTiter 96® AQueous One Solution Reagent (Promega, Madison, WI, USA) was added to each sample (20 μl per well) in a 96-well plate, and the plate was incubated at 34°C and 5% CO2. After 1 h incubation, absorbance at 490 nm was determined using a spectrophotometer (Tecan, Spark 10M, Switzerland).
2.4.3. RealTime-Glo™ MT Cell Viability assayThe RealTime-Glo™ MT Cell Viability assay (Promega, Madison, WI, USA) is a bioluminescence assay used to assess cell viability in real time. Metabolically active cells reduce the NanoLuc® luciferase prosubstrate to the luciferase substrate, thus producing a luminescence signal, which correlates with the number of viable cells. During this assay, an opaque-walled tissue culture plate was used. The 2X RealTime-Glo™ reagent was prepared with serum-free CEpiCM, and 50 μl was added into each well containing HCECs, followed by inoculation with an equal volume of serum-free medium containing amoebae in suspension at different MOIs. The plate was placed in a cell culture incubator at 34°C and 5% CO2, and cell viability was measured every 2 h. The viability was assessed in real time over 8 h, using a plate-reading luminometer (Tecan, Spark 10M, Switzerland).
2.5. MicroscopyAll cells were analyzed by phase contrast microscopy and trypan blue staining. Trypan blue facilitates the determination of the cell number and percentage of viability within a cell population (Riss et al., 2004; Strober, 2015). A μ-slide 8-well chamber (Ibidi, Martinsried, Germany) coated with collagen I was used for microscopic observation and analysis. HCECs (5 × 103 per well) were seeded in the chamber and incubated overnight. After approximately 16 h, the medium was replaced with a serum-free medium, and amoebae were added at different MOI as previously described; the plates were incubated for different time periods (2, 4, 6, 8, and 24 h). For phase contrast microscopy and microphotography, a Nikon Eclipse TE200 microscope with NIS-Elements version 4.00.07 software (Optoteam, Vienna, Austria) was used.
2.6. Statistical analysisData was analyzed through two-way ANOVA with Dunnett’s multiple comparisons test using GraphPad Prism version 7.0 for Windows (GraphPad Software Inc, San Diego, CA, USA).
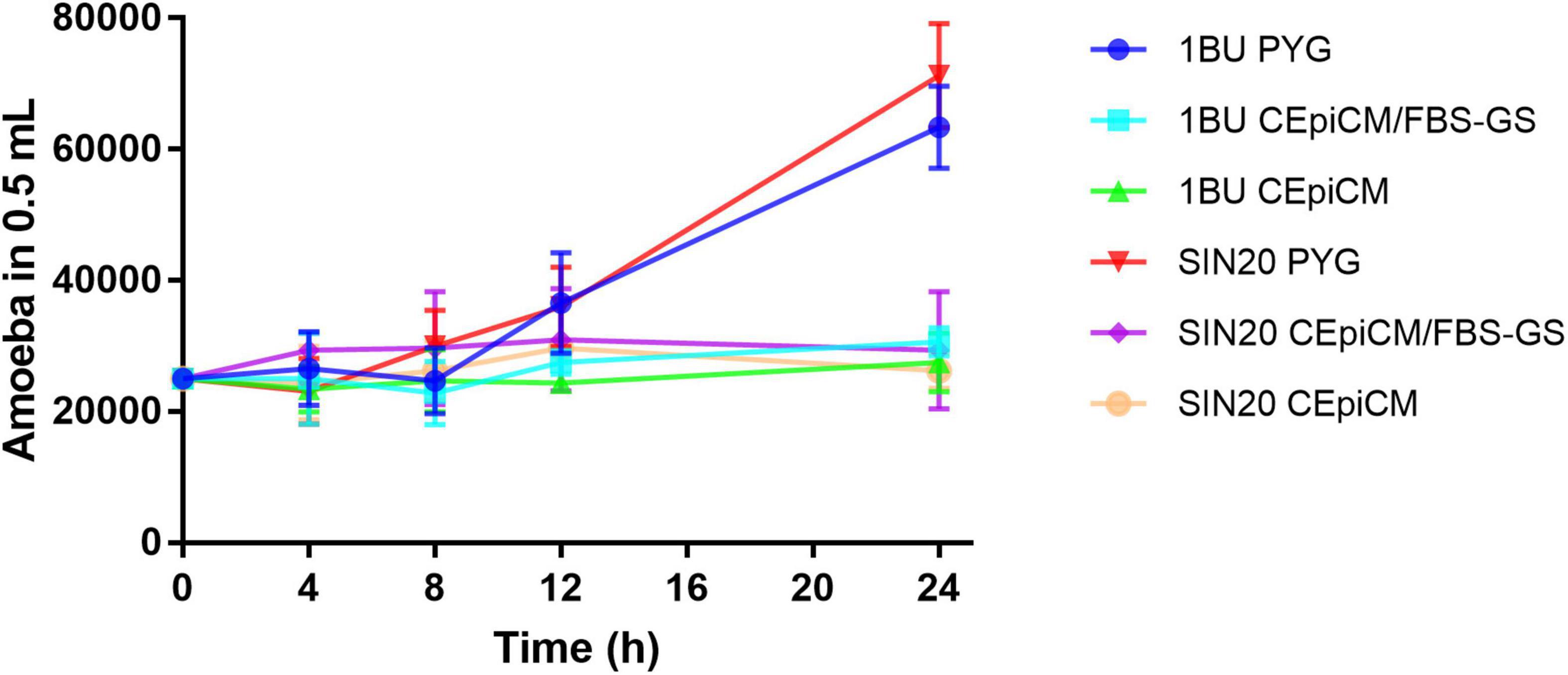
3. Results 3.1. Growth of Acanthamoeba under various conditionsTo ensure cell integrity, Acanthamoeba SIN20 and 1BU were monitored under various conditions, using CEpiCM with serum, serum-free CEpiCM, and PYG medium. Under these conditions, in serum-containing and serum-free CEpiCM, the number of amoebae on both strains remained constant over time for up to 24 h (Figure 1). Under all conditions, no dead cells were observed during the experiments. In PYG medium, the number of amoebae slightly increased after 8 h of incubation, followed by a significant increase after 12 h (Figure 1). Therefore, the time point of 8 h was selected as the maximal duration for all cytotoxicity assays, also to maintain the defined MOI ratio.
Figure 1. Growth curves of Acanthamoeba strains 1BU and SIN20 under various conditions over time at 34°C with 5% CO2 atmosphere. The numbers of amoebae were determined by cell counting with a hemocytometer.
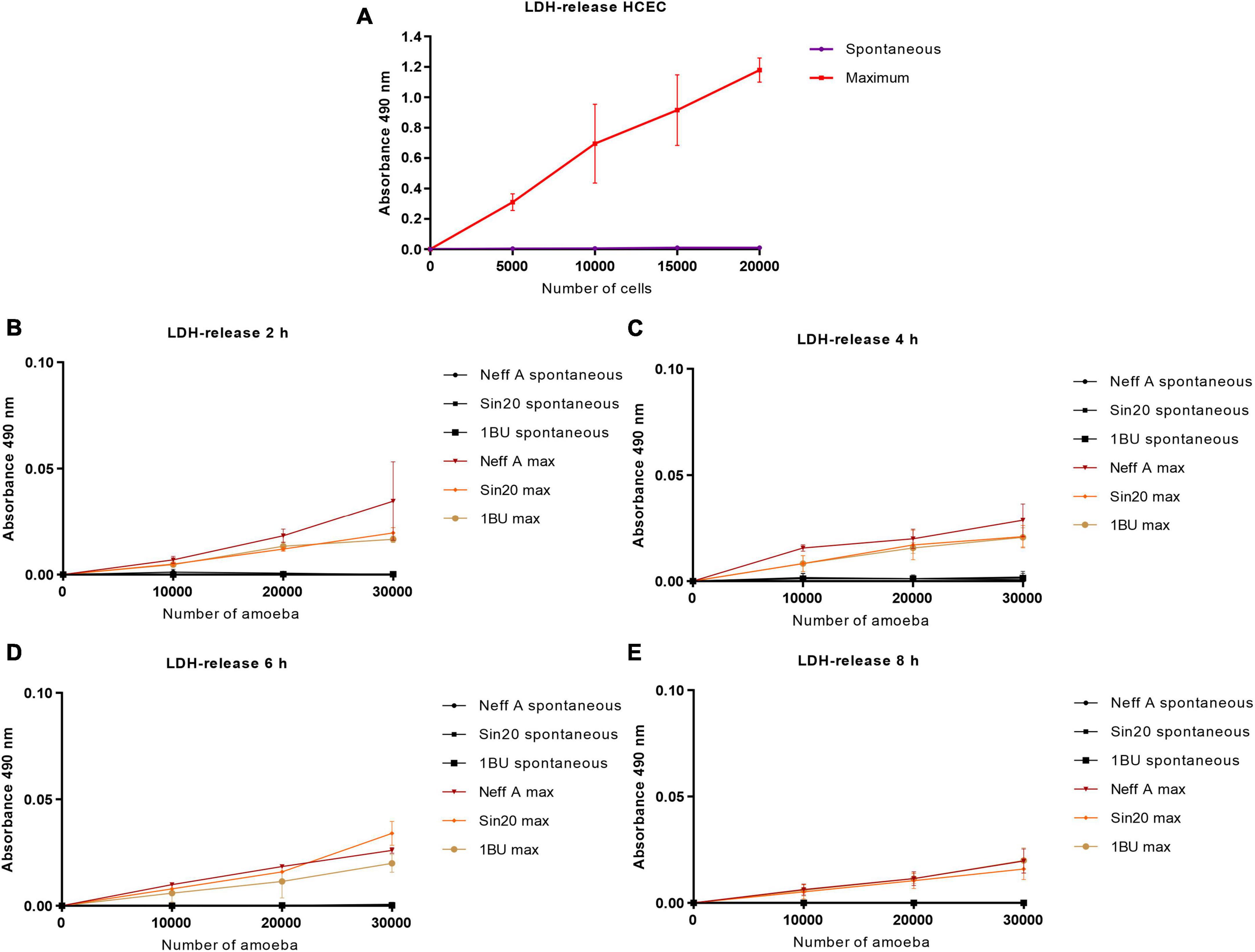
3.2. Acanthamoeba cell density affected lactate dehydrogenase productionThe effect of the cell density of the Acanthamoeba strains Neff, SIN20, and 1BU and of the HCECs on LDH activity was assessed using a defined volume of LDH lysis buffer. As shown in Figure 2, the cell density affected the linearity of LDH activity. This effect was more pronounced with HCECs (Figure 2A) than with Acanthamoeba strains (Figures 2B–E). This finding may be attributed to the fact that, owing to their larger size, mammalian cells can release more LDH into the extracellular environment than amoebae. In contrast, the incubation time and type of Acanthamoeba strain in serum-free medium did not affect LDH release.
Figure 2. Effect of HCEC (A) and amoeba (B–E) densities on absorbance at 490 nm by using the LDH release assay, at different times. The background absorbance obtained with the media alone without cells was subtracted from all data. “Maximum” corresponds to the quantity of LDH released into the cell culture medium by lysed or damaged cells, and “Spontaneous” represents the quantity released by living or non-lysed cells.
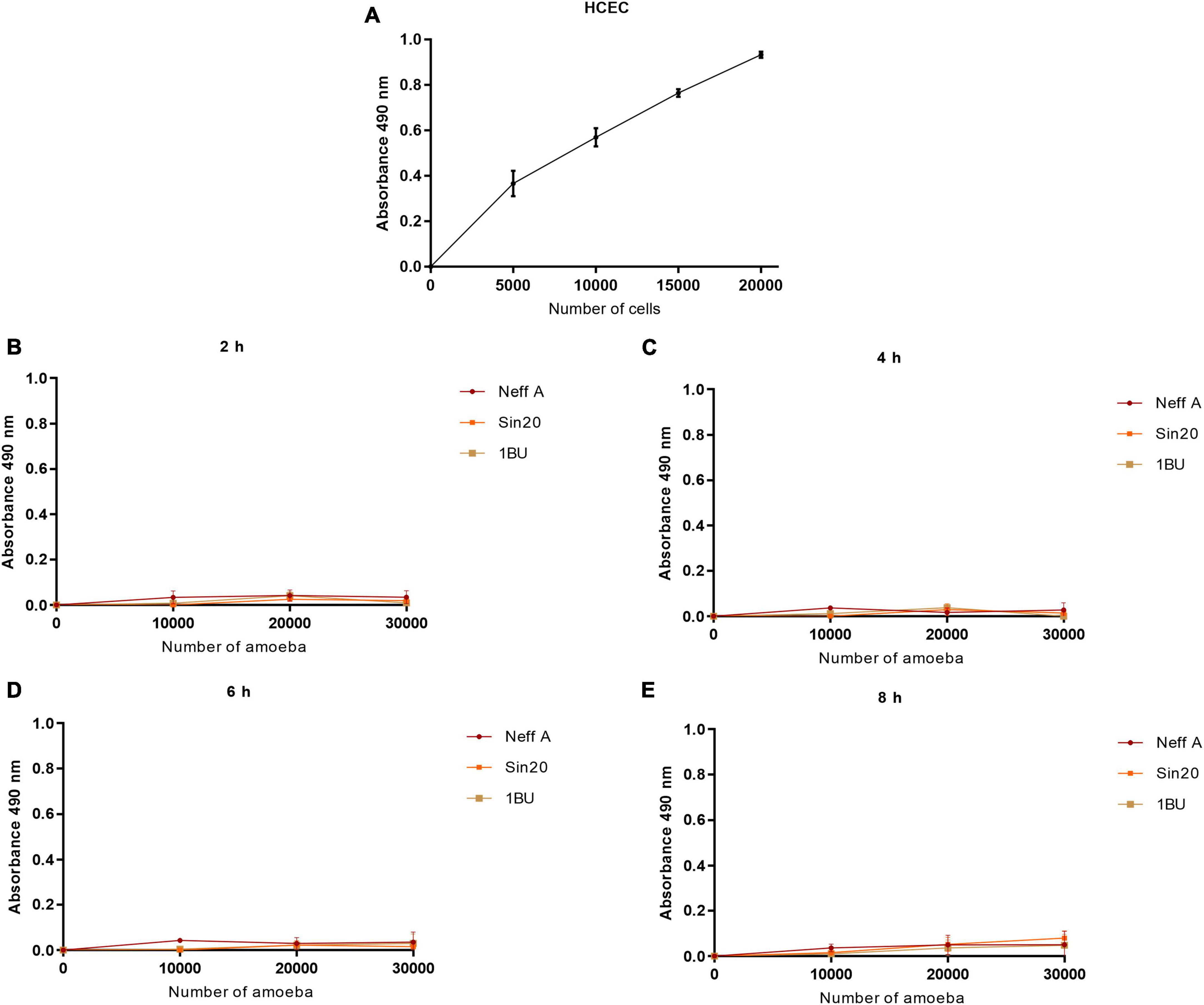
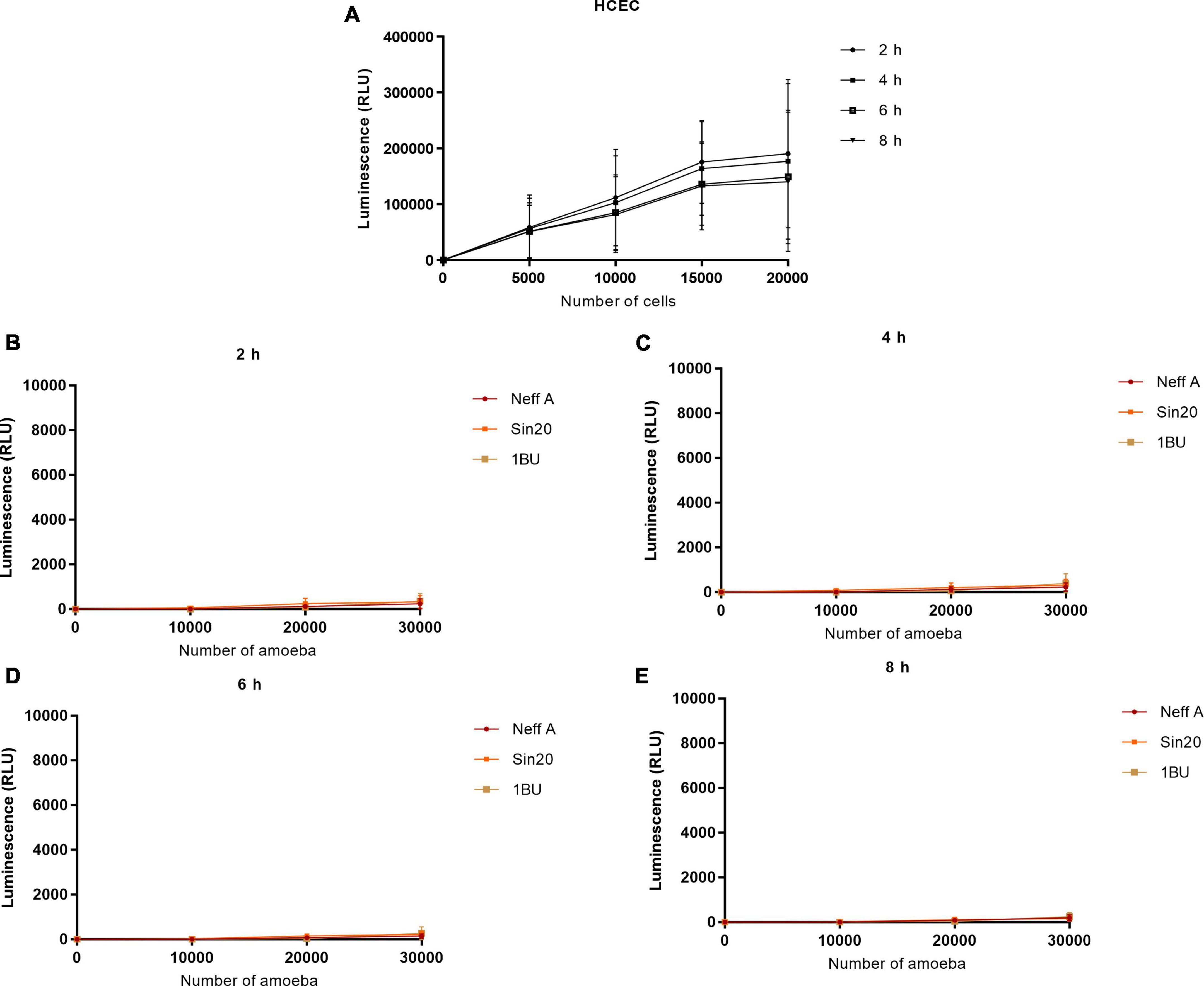
3.3. Acanthamoeba produced low signals in presence of tetrazolium salt and NanoLuc® luciferase prosubstrateSimilarly, the effect of cell number on absorbance at 490 nm and bioluminescence was evaluated using the MTS and RealTime-Glo™ MT Cell Viability assays, respectively, (Figures 3, 4). In contrast to the observations made in the LDH assay, Acanthamoeba did not increase the linear response between cell density and absorbance at 490 nm and bioluminescence, respectively, suggesting the inability of Acanthamoeba to considerably reduce the tetrazolium salt and NanoLuc® luciferase prosubstrate, respectively, and to produce a strong signal. However, a strong signal was observed with HCECs (Figures 3A, 4A). No differences in linearity were observed between the different Acanthamoeba strains used in this study.
Figure 3. Effect of HCEC (A) and amoeba (B–E) densities on absorbance at 490 nm by using the MTS assay, at different times. The background absorbance obtained with the media alone without cells was subtracted from all data.
Figure 4. Effect of HCEC (A) and amoeba (B–E) densities on the luminescence signal by using the RealTime-Glo™ MT Cell Viability assay. The background absorbance obtained with the media alone without cells was subtracted from all data.
3.4. Cytotoxic effects of Acanthamoeba on human corneal epithelial cells during co-cultureSubsequently, the cytotoxic effects of Acanthamoeba on HCECs and the effects on their viability at various incubation periods (2, 4, 6, and 8 h) and different MOIs (MOI 1, MOI 2, and MOI 3; see Table 1) were evaluated using the cytotoxicity assays. All assays were performed according to the manufacturer’s instructions, and wells containing only serum-free medium were used as controls to account for culture medium background absorbance and luminescence.
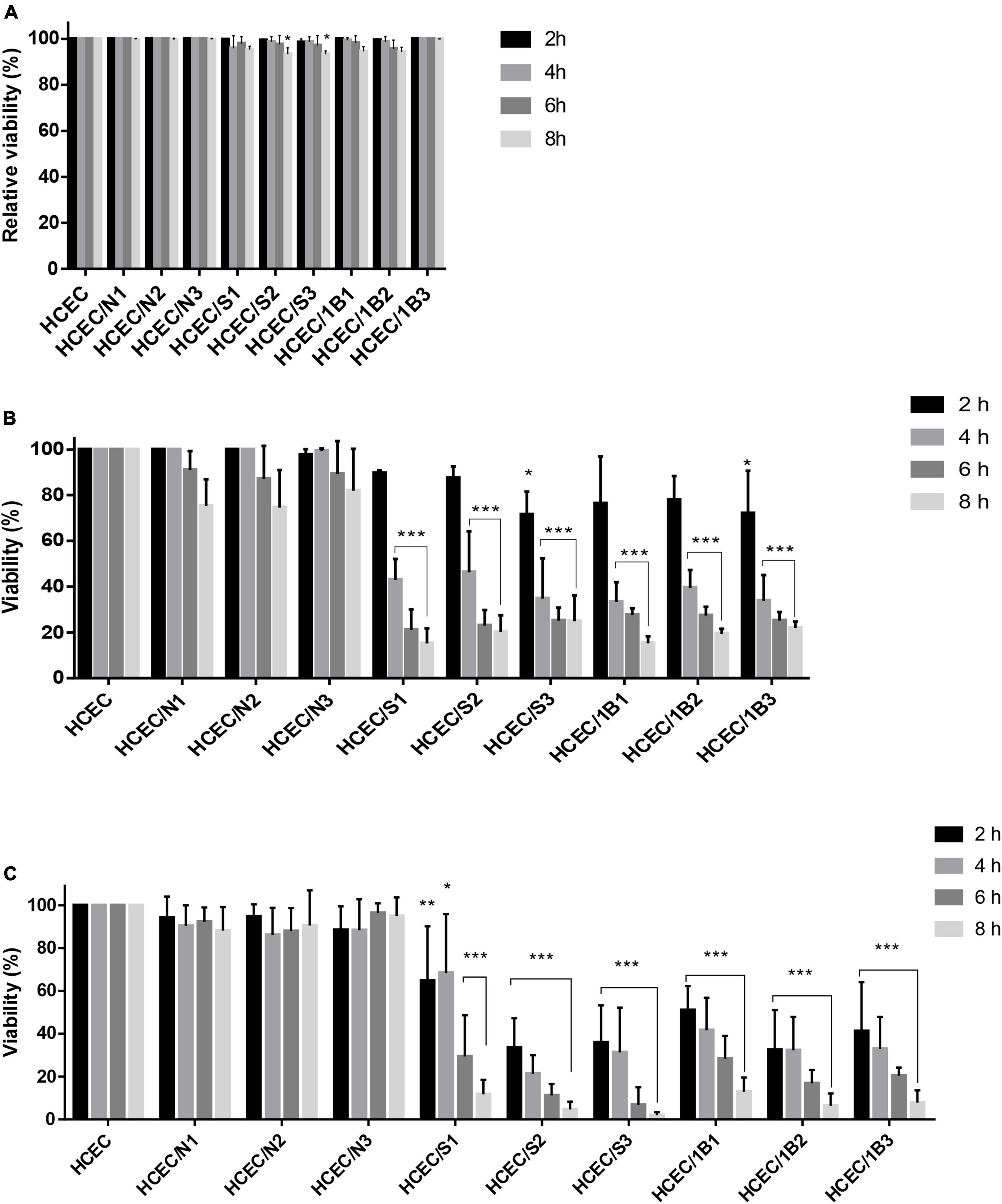
As shown in Figure 5A, in the LDH assay to evaluate Acanthamoeba-induced cytotoxicity in HCECs, no significant differences were observed in the viability of HCECs inoculated with the environmental Neff and pathogenic 1BU strains and incubated for different periods. HCECs co-cultured with the SIN20 strain showed a slight difference in the viability of the mammalian cells at MOI 2 and MOI 3 after 8 h incubation, which refers to minor cytotoxic effects of this strain, but no significant differences were observed at other MOIs and incubation times.
Figure 5. LDH release (A), MTS (B) and NanoLuc® Luciferase prosubstrate assays (C). Percentage of the viability of HCEC alone and after co-culture with Acanthamoeba strains Neff A (N), SIN20 (S) and 1BU (B) for different time periods (2, 4, 6, and 8 h), and MOIs (MOI 1: N1, S1, and B1; MOI 2: N2, S2, and B2; MOI 3: N3, S3, and B3). Values represent the means of three independent experiments, each in triplicate. Data were plotted after the correction of the media background. Statistical analysis was performed through two-way ANOVA with Dunnett’s multiple comparisons test (*P < 0.01, **P < 0.001, and ***P < 0.0001).
The MTS and RealTime-Glo™ Cell Viability assays were used to assess the effect of Acanthamoeba on the viability of HCECs in co-culture. Both pathogenic strains showed a considerable effect on the viability of HCECs in both assays, except at MOI 1 and MOI 2 after 2 h incubation in the MTS assay (Figures 5B, C). The percentage of viability of HCECs in contact with pathogenic strains 1BU and SIN20 was markedly reduced after 4, 6, and 8 h of incubation in the MTS assay and at all incubation periods in the RealTime-Glo™ Cell Viability assay (except for 1BU at MOI 1 and 4 h incubation). No significant differences were observed in the viability of HCECs in contact with the non-pathogenic Neff strain in both assays.
3.5. Microscopic examination and consistency with cytotoxicity assaysThe monolayer integrity of HCECs after inoculation with Acanthamoeba and incubation was assessed on the basis of cell detachment by phase contrast microscopy, and microphotographs were taken (Figure 6 and Supplementary material). Under all conditions, the microscopic estimation was consistent with the results obtained with the MTS and RealTime-Glo™ MT Cell Viability assays, but not with the LDH assay. The presence of cell aggregates was observed after 2 h of co-culture, and was more pronounced after 8 h of co-culture, especially at MOI 2 and MOI 3 (Figure 6). Moreover, microscopic observations after 24 h showed total or at least significant destruction of the HCEC monolayer at MOI 2 and MOI 3, while at MOI 1, numerous HCECs were still intact in co-culture (Figure 6). Owing to the difficulty in distinguishing between non-viable HCECs and non-viable Acanthamoeba SIN20, cell death could not be determined by cell counting using trypan blue staining.
Figure 6. Effect of Acanthamoeba strain SIN20 after co-culture with HCECs, at MOI 1 (B,F,J, N,R), MOI 2 (C,G,K,O,S) and MOI 3 (D,H,L,P,T) and at different times (2, 4, 6, 8, and 24 h). Panels (A,E,I,M,Q) represent the HCEC alone without co-culture. Scale bar = 50 μm. See Supplementary material for the amoeba controls.
3.6. Monitoring Acanthamoeba cytotoxicityAfter inoculation and co-culture of Acanthamoeba with HCECs, significant differences in results were observed between conventional LDH assay and other methods used in this study (Figure 5). Results of the MTS and RealTime-Glo™ MT Cell Viability assays, which were consistent with the microscopic observations of cell detachment, showed that the viability of HCECs was substantially affected by the presence of pathogenic Acanthamoeba strains, whereas almost no difference was observed with the LDH assay.
4. DiscussionThree different cytotoxicity assays, namely the LDH, MTS and RealTime-Glo™ MT Cell Viability assays, were compared in terms of their sensitivity in detecting and measuring Acanthamoeba cytotoxicity on human corneal epithelial cells (HCECs), in combination with microscopic examination. It was shown that pathogenic A. castellanii significantly reduced HCECs viability in a dose-dependent manner starting from 2 h of co-culture at varying ratios, and this effect could be monitored reliably with the MTS and RealTime-Glo™ MT Cell Viability assays, well corresponding with the observation of cell detachment and destruction.
Various methods to monitor and evaluate in vitro cell viability and cytotoxicity are available, such as colorimetric and luminescence assays and microscopic examination in combination with staining and cell counting. The methods compared in the present study, except for the rather new RealTime-Glo™ MT Cell Viability method, have been extensively used during recent years to evaluate the cytotoxicity of novel antimicrobial agents against protozoan parasites such as Leishmania, Toxoplasma gondii, Naegleria fowleri or also Acanthamoeba (Ganguly et al., 2006; Jin et al., 2009; Jha et al., 2015; López-Arencibia et al., 2015; Colon et al., 2019). The LDH assay is typically used to identify host cells that have lost membrane integrity and are considered dead on the basis of LDH release into the cell culture medium; in contrast, the MTS and RealTime-Glo™ MT Cell Viability assays use compounds that are metabolized and reduced by viable mammalian cells.
In this study, we first evaluated whether the number of amoebae (0–30,000 per well) and HCECs (0–20,000 per well) influenced the signal obtained with the LDH, MTS, and RealTime-Glo™ MT Cell Viability assays (Figures 2–4). Although HCECs produced a strong signal in all assays, this was not observed with the amoebae. While the LDH assay showed a cell density-dependent signal also for the amoebae, the MTS and RealTime-Glo™ MT Cell Viability assays showed very weak signals for various concentrations of amoebae and incubation periods, indicating that the amoebae poorly metabolized the substrates. The MTS results are consistent with a previous study, in which the density of Acanthamoeba castellanii also did not affect the tetrazolium salt reduction and produced a weak signal (Heredero-Bermejo et al., 2013). No differences in the activity of LDH released and reduction of tetrazolium salt and NanoLuc® luciferase prosubstrate were observed between the Acanthamoeba strains investigated.
Subsequently, the cytotoxic effects of the different Acanthamoeba strains on HCECs were assessed using the above-mentioned tests. Interestingly, there was a striking discrepancy between the cytotoxicity results obtained with the LDH assay and the results obtained with the two other assays as well as with the microscopic examination of cell detachment. While generally, the cytotoxicity measured with the LDH assay was similar to that reported for A. castellanii on HCECs in a previous study (Sohn et al., 2019), we found that the LDH assay underestimated the proportion of dead cells and overestimated the proportion of living cells and thus was not reliable to determine the cytotoxic effect of Acanthamoeba on HCECs. However, the LDH assay has also been used to evaluate the activity of potentially anti-amoebic agents, because this enzyme is released into the culture medium when the cell membrane integrity is affected (Sissons et al., 2005; Lorenzo-Morales et al., 2010; Anwar et al., 2018; Shi et al., 2020; Akbar et al., 2022). The results obtained in the current study corroborate the suitability of the LDH assay for evaluating Acanthamoeba viability, but not when Acanthamoeba is co-cultured with other cells. We assume that proteases produced by the pathogenic Acanthamoeba strains acted on the LDH released by the HCECs and thereby led to inaccurate results. A similar effect was reported for pathogenic bacteria in a recent study, where proteases produced by the bacteria during host–pathogen co-culture interacted with the LDH and thus lead to an underestimation of bacterial cytotoxicity (Van den Bossche et al., 2020). However, further studies will be necessary to confirm this hypothesis. In contrast to the LDH assay, the MTS and RealTime-Glo™ MT Cell Viability assays provided reliable and comparable results for Acanthamoeba cytotoxicity. The RealTime-Glo™ MT Cell Viability assay was more sensitive in determining the cytotoxic effect of Acanthamoeba; the lowest percentage of cell viability detected with the RealTime-Glo™ MT Cell Viability assay and MTS assay was approximately 2% with the SIN20 strain and 15% with the 1BU strain, respectively. Good correlation in significant findings in both pathogenic strains was observed between the two assays, especially after 4 h of co-culture with HCECs (p < 0.0001).
The ability of Acanthamoeba to establish contact with and adhere to host cells and induce apoptosis is crucial for their pathogenicity, but varies dramatically between strains. We used Acanthamoeba strains belonging to the genotype T4, since it is the most frequently genotype isolated from keratitis and non-keratitis diseases cases (Maciver et al., 2013; Walochnik et al., 2015; Castro-Artavia et al., 2017). Strains 1BU and SIN20 are AK-causing clinical isolates, isolated from patients with severe keratitis, while the Neff strain (ATCC 50373) is a non-virulent environmental isolate; and this is in line with the variations in the results obtained between 1BU and SIN20, on one hand, and Neff strains, on the other hand.
Usually, the presence of proteases and the production of extracellular vesicles are used to determine the pathogenicity of Acanthamoeba isolates (Khan et al., 2000; Walochnik and Duchêne, 2016; Moreira et al., 2020). The MTS and RealTime-Glo™ MT Cell Viability assays can be applied as valuable screening tools to assess directly the pathogenicity of Acanthamoeba isolates from environmental and clinical sources, in co-culture with mammalian cell lines.
In conclusion, the present study demonstrates the importance of choosing the most suitable method in accordance with the research purpose to meaningfully quantify cell viability and cytotoxicity in vitro. While the LDH assay, in contrast to the MTS and the RealTime-Glo™ MT Cell Viability assays, is suitable to determine Acanthamoeba viability in axenic culture, it is unsuitable to determine the cytotoxic effect of Acanthamoeba on host cells. However, the tetrazolium- and luciferase prosubstrate-based assays, although and because unsuitable to evaluate Acanthamoeba viability, were found to sensitively and reliably assess the cytotoxic effects of Acanthamoeba on human cells in vitro. To the best of our knowledge, this is the first report of a potential interference between proteases produced by Acanthamoeba with the LDH assay during Acanthamoeba–host cell co-culture leading to an underestimation of the cytotoxic effect.
Data availability statementThe original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributionsJW, ALM, and TM-P designed the study. ALM, IL-M, IH-B, RM, and TM-P collected and analyzed the data. ALM, IL-M, TM-P, and JW wrote the manuscript. All authors discussed the data, read, edited, and approved the current version of the manuscript.
FundingThis research was supported by a grant from the Austrian Science Fund (P 35259).
AcknowledgmentsWe thank the members of the unit Molecular Parasitology at the Institute of Specific Prophylaxis and Tropical Medicine of the Medical University of Vienna for their technical support.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1175469/full#supplementary-material
ReferencesAkbar, N., El-Gamal, M., Saeed, B., Oh, C., Abdel-Maksoud, M., Khan, N., et al. (2022). Antiamoebic activity of imidazothiazole derivatives against opportunistic pathogen Acanthamoeba castellanii. Antibiotics 11, 1–11.
PubMed Abstract | Google Scholar
Anwar, A., Siddiqui, R., Hussain, M., Ahmed, D., Shah, M., and Khan, N. (2018). Silver nanoparticle conjugation affects antiacanthamoebic activities of amphotericin B, nystatin, and fluconazole. Parasitol. Res. 117, 265–271. doi: 10.1007/s00436-017-5701-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Castro-Artavia, E., Retana-Moreira, L., Lorenzo-Morales, J., and Abrahams-Sandí, E. (2017). Potentially pathogenic Acanthamoeba genotype T4 isolated from dental units and emergency combination showers. Mem. Inst. Oswaldo Cruz. 112, 817–821. doi: 10.1590/0074-02760170147
PubMed Abstract | CrossRef Full Text | Google Scholar
Clarke, D., and Niederkorn, J. (2006). The pathophysiology of Acanthamoeba keratitis. Trends Parasitol. 22, 175–180.
Colon, B., Rice, C., Guy, R., and Kyle, D. (2019). Phenotypic screens reveal posaconazole as a rapidly acting amebicidal combination partner for treatment of primary amoebic meningoencephalitis. J. Infect. Dis. 219, 1095–1103. doi: 10.1093/infdis/jiy622
PubMed Abstract | CrossRef Full Text | Google Scholar
Ganguly, S., Bandyopadhyay, S., Sarkar, A., and Chatterjee, M. (2006). Development of a semi-automated colorimetric assay for screening anti-leishmanial agents. J. Microbiol. Methods 66, 79–86. doi: 10.1016/j.mimet.2005.10.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Garate, M., Cao, Z., Bateman, E., and Panjwani, N. (2004). Cloning and characterization of a novel mannose-binding protein of Acanthamoeba. J. Biol. Chem. 279, 29849–29856. doi: 10.1074/jbc.M402334200
PubMed Abstract | CrossRef Full Text | Google Scholar
Heredero-Bermejo, I., Copa-Patiño, J., Soliveri, J., Gómez, R., De La Mata, F., and Pérez-Serrano, J. (2013). In vitro comparative assessment of different viability assays in Acanthamoeba castellanii and Acanthamoeba polyphaga trophozoites. Parasitol. Res. 112, 4087–4095. doi: 10.1007/s00436-013-3599-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Jha, B., Jung, H., Seo, I., Suh, S., Suh, M., and Baek, W. (2015). Juglone induces cell death of Acanthamoeba through increased production of reactive oxygen species. Exp. Parasitol. 159, 100–106. doi: 10.1016/j.exppara.2015.09.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Jin, C., Kaewintajuk, K., Jiang, J., Jeong, W., Kamata, M., Kim, H., et al. (2009). Toxoplasma gondii: a simple high-throughput assay for drug screening in vitro. Exp. Parasitol. 121, 132–136.
Khan, N. (2006). Acanthamoeba: biology and increasing importance in human health. FEMS Microbiol. Rev. 30, 564–595.
Khan, N., Jarroll, E., Panjwani, N., Cao, Z., and Paget, T. (2000). Proteases as markers for differentiation of pathogenic and nonpathogenic species of Acanthamoeba. J. Clin. Microbiol. 38, 2858–2861. doi: 10.1128/JCM.38.8.2858-2861.2000
PubMed Abstract | CrossRef Full Text | Google Scholar
Koehsler, M., Leitsch, D., Duchêne, M., Nagl, M., and Walochnik, J. (2009). Acanthamoeba castellanii: growth on human cell layers reactivates attenuated properties after prolonged axenic culture. FEMS Microbiol. Lett. 299, 121–127. doi: 10.1111/j.1574-6968.2009.01680.x
PubMed Abstract | CrossRef Full Text | Google Scholar
List, W., Glatz, W., Riedl, R., Mossboeck, G., Steinwender, G., and Wedrich, A. (2021). Evaluation of Acanthamoeba keratitis cases in a tertiary medical care centre over 21 years. Sci. Rep. 11, 1–10. doi: 10.1038/s41598-020-80222-3
PubMed Abstract | CrossRef Full Text | Google Scholar
López-Arencibia, A., García-Velázquez, D., Martín-Navarro, C., Sifaoui, I., Reyes-Batlle, M., Lorenzo-Morales, J., et al. (2015). In vitro activities of hexaazatrinaphthylenes against Leishmania spp. Antimicrob. Agents Chemother. 59, 2867–2874.
Lorenzo-Morales, J., Khan, N. A., and Walochnik, J. (2015). An update on Acanthamoeba keratitis: Diagnosis, pathogenesis and treatment. Parasite, Vol. 22. Paris, 10. doi: 10.1051/parasite/2015010
PubMed Abstract | CrossRef Full Text | Google Scholar
Lorenzo-Morales, J., Martín-Navarro, C., López-Arencibia, A., Santana-Morales, M., Afonso-Lehmann, R., Maciver, S., et al. (2010). Therapeutic potential of a combination of two gene-specific small interfering RNAs against clinical strains of Acanthamoeba. Antimicrob. Agents Chemother. 54, 5151–5155. doi: 10.1128/AAC.00329-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Loufouma Mbouaka, A., Leitsch, D., Koehsler, M., and Walochnik, J. (2021). Antimicrobial effect of auranofin against Acanthamoeba spp. Int. J. Antimicrob. Agents. 58:106425.
Maciver, S., Asif, M., Simmen, M., and Lorenzo-Morales, J. (2013). A systematic analysis of Acanthamoeba genotype frequency correlated with source and pathogenicity: T4 is confirmed as a pathogen-rich genotype. Eur. J. Protistol. 49, 217–221. doi: 10.1016/j.ejop.2012.11.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattana, A., Cappai, V., Alberti, L., Serra, C., Fiori, P., and Cappuccinelli, P. (2002). ADP and other metabolites released from Acanthamoeba castellanii lead to human monocytic cell death through apoptosis and stimulate the secretion of proinflammatory cytokines. Infect. Immun. 70, 4424–4432. doi: 10.1128/IAI.70.8.4424-4432.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Mazur, T., and Hadaś, E. (1994). The effect of the passages of Acanthamoeba strains through mice tissue on their virulence and its biochemical markers. Parasitol. Res. 80, 431–434. doi: 10.1007/BF00932382
PubMed Abstract | CrossRef Full Text | Google Scholar
Moreira, L., Ramírez, D., Linares, F., Ledezma, A., Garro, A., Osuna, A., et al. (2020). Isolation of Acanthamoeba t5 from water: characterization of its pathogenic potential, including the production of extracellular vesicles. Pathogens 9:144. doi: 10.3390/pathogens9020144
PubMed Abstract | CrossRef Full Text | Google Scholar
Morrison, A., Morris, R., Shannon, A., Lauer, S., Guarner, J., and Kraft, C. (2016). Disseminated Acanthamoeba infection presenting with cutaneous lesions in an immunocompromised patient: a case report, review of histomorphologic findings, and potential diagnostic pitfalls. Am. J. Clin. Pathol. 145, 266–270. doi: 10.1093/ajcp/aqv081
PubMed Abstract | CrossRef Full Text | Google Scholar
Randag, A., Van Rooij, J., Van Goor, A., Verkerk, S., Wisse, R., Saelens, I., et al. (2019). The rising incidence of Acanthamoeba keratitis: a 7-year nationwide survey and clinical assessment of risk factors and functional outcomes. PLoS One 14:e0222092. doi: 10.1371/journal.pone.0222092
PubMed Abstract | CrossRef Full Text | Google Scholar
Riss, T., Moravec, R., Niles, A., Duellman, S., Benink, H., Worzella, T., et al. (2004). Cell viability assays. Assay Guid. Man. 1–25. Available online at: http://www.ncbi.nlm.nih.gov/pubmed/23805433 (accessed October 2022).
Shi, L., Muthukumar, V., Stachon, T., Latta, L., Elhawy, M., Gunaratnam, G., et al. (2020). The effect of anti-amoebic agents and Ce6-PDT on Acanthamoeba castellanii trophozoites and cysts, in vitro. Transl. Vis. Sci. Technol. 9, 1–15. doi: 10.1167/tvst.9.12.29
PubMed Abstract | CrossRef Full Text | Google Scholar
Siddiqui, R., Chaudhry, T., Lakhundi, S., Ahmad, K., and Khan, N. (2014). Failure of chemotherapy in the first reported cases of Acanthamoeba keratitis in Pakistan. Pathog. Glob. Health. 108, 49–52. doi: 10.1179/2047773213Y.0000000124
PubMed Abstract | CrossRef Full Text | Google Scholar
Sissons, J., Kwang, S., Stins, M., Jayasekera, S., Alsam, S., and Khan, N. (2005). Acanthamoeba castellanii induces host cell death via a phosphatidylinositol 3-kinase-dependent mechanism. Infect. Immun. 73, 2704–2708. doi: 10.1128/IAI.73.5.2704-2708.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Sohn, H., Seo, G., Lee, J., Ham, A., Oh, Y., Kang, H., et al. (2019). Cytopathic change and inflammatory response of human corneal epithelial cells induced by Acanthamoeba castellanii trophozoites and cysts. Korean J. Parasitol. 57, 217–223. doi: 10.3347/kjp.2019.57.3.217
PubMed Abstract | CrossRef Full Text | Google Scholar
Strober, W. (2015). Trypan blue exclusion test of cell viability. Curr Protoc Immunol 111, A3.B.1–A3.B.3.
Van den Bossche, S., Vandeplassche, E., Ostyn, L., Coenye, T., and Crabbé, A. (2020). Bacterial interference with lactate dehydrogenase assay leads to an underestimation of cytotoxicity. Front. Cell Infect. Microbiol. 10:494. doi: 10.3389/fcimb.2020.00494
PubMed Abstract | CrossRef Full Text | Google Scholar
Walochnik, J., and Duchêne, M. (eds). (2016). Molecular parasitology: Protozoan parasites and their molecules, Vol. IX. Vienna: Springer Publishing, 1–547.
留言 (0)