
記住我
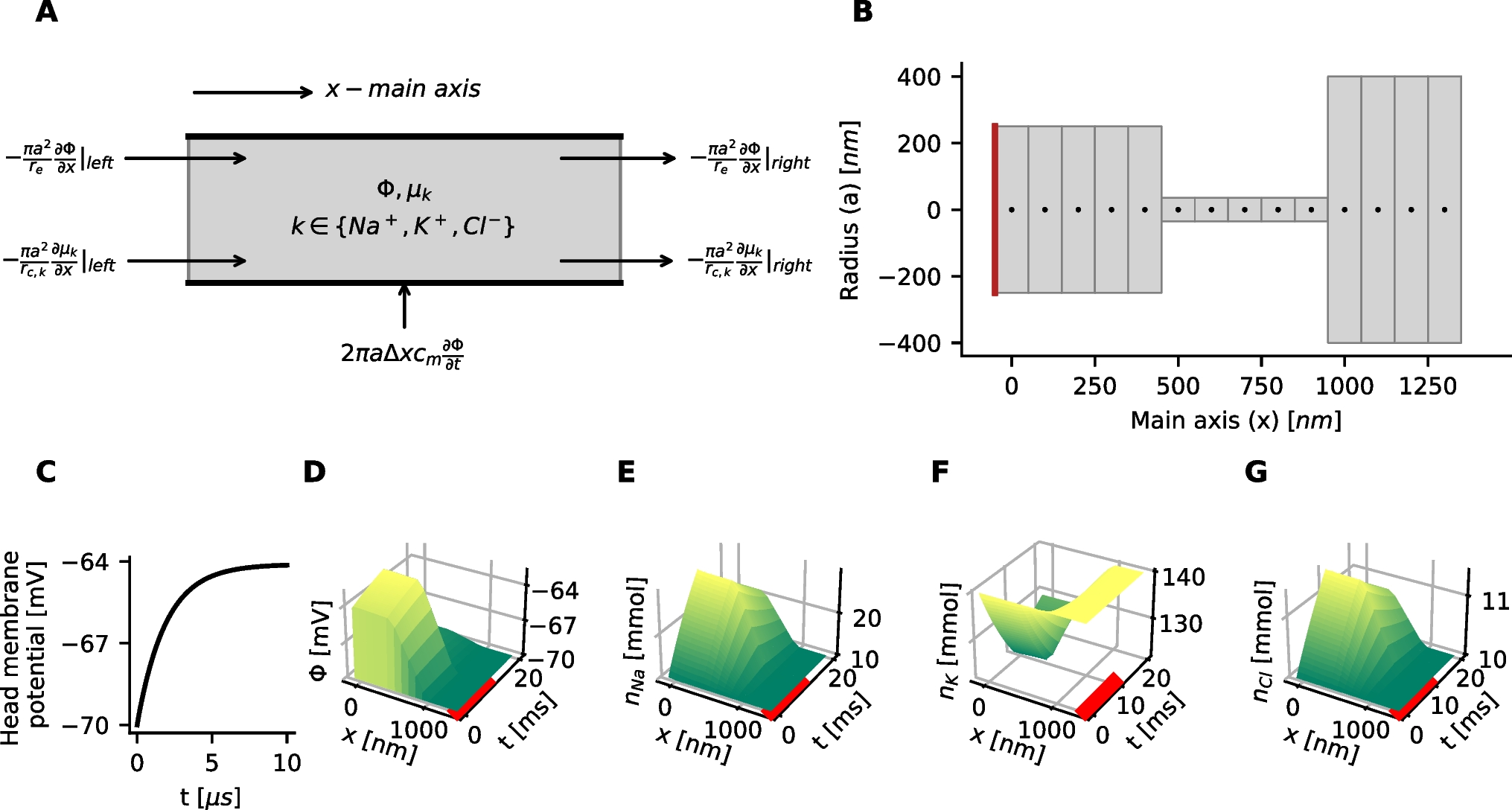



We studied how \(f-I\) curves in the nine models described in the methods section were sensitive to a selection of model parameters, including the specific membrane capacitance \(c_\text \), maximal conductances \(\bar_X\) for various ion channels, and ionic reversal potentials \(E_k\).
3.1 Effects of \(c_\text \) on firing propertiesThe firing properties of all models were sensitive to the value of \(c_\text /c_\text \), where \(c_\text \) is the model’s default value of the capacitance. An example illustration is given in Fig. 2A showing the voltage trace of Allen model 1, where an increasing \(c_\text \) lead to a broadening of the spikes, a lower spike amplitude and a decreased firing rate. The broadening was general for all models (Fig. 2E, F), as expected, since \(c_\text \propto \tau _\text \) should slow down the membrane dynamics.
Fig. 2
Properties of the neuron model as a function of specific membrane capacitance \(c_\text \). \(c_\text 0\) is the default value of \(c_\text \) in the model. A Voltage trace for Allen model 1 for default \(c_\text \) and \(1.5c_\text \). Inset: normalized voltage trace over the duration of one peak. The two traces have been shifted to align the peak maxima, B Threshold current vs \(c_\text \), C Frequency f vs \(c_\text \) for all models for input current I = 0.2 nA, D f vs \(c_\text \) for the Allen models for I = 0.4 nA, E Spike duration (defined as the width of the spike at -40 mV) for I = 0.2 nA, F Spike duration at -40 mV for I = 0.4 nA. Note that the one-compartment model and the ball-and-stick model do not fire for \(I=0.4\) nA. OC - one-compartment model, BAS - ball-and-stick model, A1 - Allen model 1, A2 - Allen model 2, A3 - Allen model 3, all - \(c_\text \) changed at every segment of the neuron, sprx - \(c_\text \) only changed at the soma and proximal dendrites
Note that the Allen models exhibit narrower spikes than the HH models, resembling the short spike duration observed in PV cells (Bartos & Elgueta, 2012).
The effect of \(c_\text \) on the threshold current for firing onset is shown in Fig. 2B. The threshold current remained constant or varied only slightly with the capacitance, except for the ball-and-stick model. However, in none of the models the onset was shifted when varying \(c_\text \) over the interval relevant under Tewari et al.’s experiments (i.e. from default to a factor 1.5 increase). This held both for simulations where \(c_\text \) had only been altered in the soma and proximal dendrites, as indicated by dashed lines, and simulations where \(c_\text \) was changed everywhere, as indicated by solid lines.
There was no clear general trend shared among the models in terms of how the firing frequency f depended on \(c_\text \). Over the same \(c_\text \) interval, f increased with \(c_\text \) in some models, while it decreased with \(c_\text \) in others (Fig. 2C, D). This was also the case for the \(c_\text \) interval relevant under the experiments by Tewari et al. However, for the strongest of the current injections considered (0.4 nA in Fig. 2D), all models except Allen model 2 (which stopped firing at \(\sim\) 25% increase in \(c_\text \)) showed a decreasing trend in f with \(c_\text \). This suggests that at least the maximal firing rate in these models should be reduced, like in the experiments, when \(c_\text \) is increased (as an effect of PNN degradation).
For all the Allen models, f vs \(c_\text \) varied less when \(c_\text \) was only changed in the soma and proximal dendrites, which is to be expected as we altered \(c_\text \) on a smaller part of the neuron. For Allen model 3, for instance, this graph appeared far less curved when \(c_\text \) was changed in the soma and the proximal dendrites compared to when \(c_\text \) was changed everywhere. However, within the range \(c_\text /c_\text \in [1.0,1.5]\), the difference between the two cases was relatively small. In the following, we therefore show results only for the supposedly more realistic case where PNN degradation is assumed to alter \(c_\text \) only on the soma and proximal dendrites (results for \(c_\text \) altered everywhere is found in Supplementary Fig. 1).
Fig. 3
Frequency-input curves for selected values of \(c_\text \) for the various models. \(c_\text \) is altered in the soma and proximal dendrites. A The one-compartment Hodgkin-Huxley model, B The ball-and-stick Hodgkin-Huxley model, C Allen model 1, D Allen model 2, E Allen model 3, F The relative difference in f between the \(1.0c_\text \)- and \(1.5c_\text \) curves computed at the largest current that gave sustained firing in both cases
To compare with the \(f-I\) curves of Tewari et al. (2018) (Fig. 1) we stimulated the different neuron models with a range of input currents for various values of \(c_\text \). Except for stimuli near the onset threshold, all models displayed a reduction in f when increasing \(c_\text \) (Fig. 3).
In addition to affecting the firing rate, changes in \(c_\text \) caused a shift in the spiking onset threshold in some of the models (Fig. 3). However, in none of the models the onset was shifted when varying \(c_\text \) over the interval relevant under the Tewari et al.’s experiments.
The HH models (Fig. 3A, B) exhibited type II firing, meaning that the firing rate increases abruptly from zero to a higher value when the threshold current is reached. However, Tewari et al. observed Type I firing in their experiments. The HH models are therefore not ideal for simulating PV cells.
The Allen models, which were constructed based on morphological and electrophysiological recordings from real PV cells, had \(f-I\) curves that were more similar to the the experimental recordings. For Allen model 1 the firing frequency increased with \(c_\text \) for input currents close to the threshold current (Fig. 3C). At an input current of around \(I=0.28\) nA, the \(f-I\) curves crossed, after which the firing frequency decreased with \(c_\text \). For Allen model 2, the \(f-I\) curve crossings started closer to the threshold current and were less pronounced (Fig. 3D). For a relatively larger range of input currents f decreased with increasing \(c_\text \). For Allen model 3, the firing frequency was approximately equal for all \(c_\text \) for stimuli up to \(I=0.18\) nA, after which f started to decrease as \(c_\text \) was increased (Fig. 3E).
None of the \(f-I\) curves in Fig. 3 show a sufficient reduction in firing when reducing \(c_\text \) by 25 or 50% to explain the observations in the experiments by Tewari et al. (as seen from Fig. 3G). In other words, changing \(c_\text \) was on its own not enough to reproduce their findings. We therefore hypothesized that PNN degradation affected additional mechanisms which also contributed to the observed reduction in f. It has been reported that PNNs might affect both local concentrations of ions (Morawski et al., 2015; Burket et al., 2021) or currents through ion channels (Vigetti et al., 2008; van ’t Spijker & Kwok, 2017). In the following sections, we have therefore explored how variations in reversal potentials \(E_k\) and ion channel conductances \(\bar_X\) affect the firing frequency of our model neurons.
As the HH models contained relatively few of the membrane mechanisms present in PV neurons, and also produced type II firing unlike the type I firing seen in Tewari et al.’s experiments, we excluded them from our further analyses, and focused on the Allen models.
3.2 Effects of reversal potentials on firing ratesPNNs have been shown to be involved in the regulation of ionic concentrations (Morawski et al., 2015; Burket et al., 2017), and it is therefore likely that PNN degradation will lead to changes in ionic reversal potentials. This may in turn have dramatic consequences for neural firing properties, as has been the topic of many previous studies (Kager et al., 2000; Wei et al., 2014; Sætra et al., 2020). In order to gauge their general effect on the firing in PV neurons, all reversal potentials in the Allen models were changed separately by up to \(\pm 20\) mV, as shown in Fig. 4.
Fig. 4
Frequency-input curves when varying the different reversal potentials in the Allen models. Note that the reversal potential of calcium in the Allen models was found using calcium dynamics together with Eq. (4), so \(E_}\) is given at \(t=0\) ms and will vary throughout the simulations
The simulations suggested that, among the reversal potentials, \(E_\text \) (all Allen models: Fig. 4A-C) and \(E_\text \) (in Allen model 1: Fig. 4J) seemed the most likely candidates to have contributed during the experiments by Tewari et al. Both these led to moderate changes in the firing rate without strongly affecting the onset of firing. In contrast, changes in \(E_\text \) and \(E_\text \) (Fig. 4D-I) caused large shifts in firing onset not seen in the experiments, or no effect at all (Fig. 4H).
3.3 Effect of conductances on firing ratesTo gauge the effect of conductance changes on \(f-I\) curves, we varied all conductances one-by-one over an interval ranging from 0.3\(\bar_X\) to 10.0\(\bar_X\), where \(\bar_X\) is the default maximal conductance (for fully open ion channels). Among the nine models, Allen model 1 responded most strongly to conductance changes. We therefore show results only for that model (Figure 5). The effect of conductance on Allen models 2 and 3 is shown in (Supplementary Figs. 2 and 3).
Fig. 5
Frequency-input curves when varying different conductances in Allen model 1. \(\bar_X\) is the default value of the conductance. A \(\bar_}\), B \(\bar_}\), C \(\bar_}\), D \(\bar_}\), E \(\bar_}\), F \(\bar_}\), G \(\bar_}\), H \(\bar_}\), I \(\bar_}\), J \(\bar_}\), K \(\bar_}\)
3.3.1 Calcium conductancesIn many neuron types, inward depolarizing Ca\(^\) currents trigger outward hyperpolarizing K\(^\) currents through Ca\(^\)-activated K\(^\) channels (see e.g. Destexhe & Sejnowski (2003) or Halnes et al. (2011)). Hence, whether the overall effect of a Ca\(^\) current leads to an increased or decreased firing rate generally depends on the neuron’s ion channel composition.
In Allen model 1, the direct depolarizing effect associated with inward Ca\(^\) currents was much smaller than the secondary hyperpolarizing effects associated with the activation of Ca\(^\)- activated SK channels. Increasing \(\bar_}\) thus had a negative effect on the firing rate in this model (Fig. 5A). Increasing \(\bar_}\) by factors 3, 7 and 10, lead to quite pronounced decreases in f by 28%, 54% and 65%, respectively, at the maximal current injection considered (0.8 pA). In comparison, the decrease in f (at the maximal current injection) in Tewari et al.’s experiments was 38%. The increased conductance did not lead to a shift in the onset of firing. Likewise, reductions in f (without a shift in the onset threshold) could also be obtained by an increase in \(\bar_}\) (Fig. 5B).
Combined with changes in \(c_\text \) and possibly other mechanisms, \(\bar_}\) and \(\bar_}\) could be candidate mechanisms for explaining effects of PNN degradation on firing properties. However, we did not find experimental studies in support of the notion that PNN degradation should increase Ca\(^\) conductances. Contrarily, in retinal photoreceptors, chondroitin sulfates, which are key components of the PNNs, were found to shift the activation curve of unspecified calcium channels towards lower voltages (Vigetti et al., 2008). Hence, if removing PNNs means removing chondroitin sulfates, we would expect activation to shift towards higher values, resulting in generally reduced calcium current \(I_}\). Likewise, in experiments on hippocampal slices, (Kochlamazashvili et al., 2010) found that \(I_}\) was reduced upon breakdown of PNN component hyaluronan by hyaluronidase, and increased when hyaluronan was added to the hyaluronidase-treated neurons. If anything, the cited experiments thus suggest that PNN degradation should decrease overall calcium currents, rather than increase them, as we needed to do to reduce f in Allen model 1. Hence, we do no not consider \(\bar_}\) or \(\bar_}\) as main candidates for explaining Tewari et al.’s results.
We note that while \(\bar_}\) had almost no effect on f in Allen models 2 and 3 (Supplementary Figs. 2A and 3A), increases in \(\bar_}\) had a small positive effect on f in these models (Supplementary Figs. 2B and 3B). The latter suggests that in these models, the depolarizing effect of \(I_}\) dominated over indirect hyperpolarizing effects via SK activation. However, f was insensitive to reductions in \(\bar_}\) and \(\bar_}\) in these models. Hence, the decrease in f observed in Tewari et al.’s experiments could not be obtained by reducing Ca\(^\) conductances in any of the Allen models.
3.3.2 Sodium conductanceAn increase in \(\bar_}\) lead to a downward (towards lower input) shift in the onset of firing (Fig. 5C) in Allen model 1, and thus and increased f for weak stimuli. However, the \(f-I\) curves for various \(\bar_}\) crossed at about \(I=0.22\) nA, and for input stronger than this, increase in \(\bar_}\) caused a decrease in f, as has been seen in a previous modeling study (Kispersky et al., 2012).
There is experimental support that PNNs affect NaV currents. Tenascin-C and net component tenascin-R have been found to play a crucial role in localizing NaV channels in the axon initial segment and nodes of Ranvier (Srinivasan et al., 1998), and tenascin-R has also been found to increase the maximum amplitude of NaV currents when in solution, thus indicating an increase in \(\bar_}\) (Xiao et al., 1999).
As tenascin-R is a crosslinker in the nets, it is unclear whether it would get close enough to the NaV channels to affect them when present in intact PNNs. It is possible that removing the nets would lead to free tenascin-R and hence increased \(\bar_}\), but this effect might be transient due to diffusion of tenascin-R away from the cell surface. If tenascin-R lingers near the cell membrane after dissolving PNNs, a resulting increase in \(\bar_}\) could, as we saw in Fig. 5C, partially explain the decrease in firing in Fig. 1. However, increases in \(\bar_}\) produced pronounced shifts in the onset of firing not seen in the Tewari et al.’s experiments, and changes in \(\bar_}\) thus does not seem like a main candidate for explaining the experiments.
3.3.3 Potassium conductance: \(\bar_}\)Ion channel Kv3.1b is often highly expressed in PV neurons, which are often enwrapped in PNNs (Favuzzi et al., 2017). Experiments have also suggested that PNNs affect Kv3.1b channels. In brevican knock-out mice, clustering of these channels were altered, and active Kv3.1b was increased (Favuzzi et al., 2017). As PNNs contain brevican, it thus seems natural to expect that PNN degradation should lead to an increase in Kv3.1b conductance and hence \(\bar_}\).
The above evidence suggests that effects of PNN on \(\bar_}\) could be an important contributor to the reduction in f seen in Fig. 1. However, increasing \(\bar_}\) only gave a small reduction in f, but a pronounced shift towards higher input in the onset of firing (Fig. 5D), not seen in Tewari et al.’s experiments. According to the simulations, \(\bar_}\) is thus not a good candidate mechanism for explaining Tewari et al.’s experiments.
3.3.4 Potassium conductance: \(\bar_}\) and \(\bar_}\)Moderate and quite similar reductions in f could be obtained by increasing \(\bar_}\) (Fig. 5E) and \(\bar_}\) (Fig. 5F). Neither of these mechanisms affected the onset of the \(f-I\) curve significantly. The increase in \(\bar_}\) has experimental support, as attenuation of the extracellular matrix through application of chondroitinase ABC have been shown to upregulate SK-channels in hippocampal neurons, leading to an increase in \(I_}\) by, on average, a factor 3 (see Fig. 2f in Dembitskaya et al. (2021)). When it comes to \(\bar_}\), we found no mention in the literature as to whether it is affected by PNNs. As the curves look promising and the literature does not exclude them, we consider both these conductances as candidate mechanisms for explaining parts of the reduction in f found in the experiments of Tewari et al.
3.3.5 Other potassium conductancesThe K\(^+\) conductances \(\bar_}\) (Fig. 5G) and \(\bar_}\) (Fig. 5H) had little impact on f. Also, we have not found any mentions in the experimental literature suggesting that PNN affect these currents, and do not consider them as candidates for explaining Tewari et al.’s experiments.
In contrast, \(\bar_}\) induced a clear shift in the onset of firing, as seen from Fig. 5I. Its \(f-I\) curves (for various values of \(\bar_}\)) crossed at different input currents. For low input currents, f decreased with increasing \(\bar_}\), while for larger input currents f increased with decreasing \(\bar_}\). Due to the relatively large shifts and lack of mention in the literature, we do not consider \(\bar_}\) as a main candidate for explaining Tewari et al.’s experiments.
3.3.6 \(\bar_}\)The hyperpolarization activated \(I_\text \) current was almost inactive during the depolarizing current injections used in our simulations, and presumably also in Tewari et al.’s experiments. Changing \(\bar_}\) in Allen model 1 thus had almost no impact on its \(f-I\) curves (Fig. 5J). Due to its low impact on the firing frequency, we conclude that \(\bar_}\) is not a candidate mechanism for explaining the reduction in f found in Fig. 1. We note that chondroitin sulfates, which are present in PNNs have been found to shift the activation curve of \(I_\text \) in photoreceptors (Vigetti et al., 2008), but PNNs were not found to have any effect on \(I_\text \) in deep cerebellar nuclei (Hirono et al., 2018). As previously explained, we have focused on conductances of various channels, and have not tried to account for activation kinetics.
3.3.7 Leak conductanceA decrease in f could also be obtained by increasing the leak conductance \(\bar_}\) (Fig. 5K). However, similarly to \(\bar_}\), \(\bar_}\) induced a clear shift in the onset of firing. Also, changes in the membrane resistance consistent with a change in \(\bar_}\) were not found in the experiments by Tewari et al. (2018). We therefore do not consider \(\bar_}\) as a main candidate for explaining Fig. 1.
3.4 A combinatorial explanationIn the experiments by Tewari et al. (2018), PNN degradation lead to a maximum reduction in \(c_\text \) by 50%. As the simulations in Fig. 3G suggested, such a change in \(c_\text \) did reduce the firing rate in fast-spiking interneuron, but not sufficiently to explain the experiments in Fig. 3G). The parameter explorations in Sections 3.2 and 3.3 allowed us to identify possible candidate mechanisms that, combined with the observed change in \(c_\text \), could explain the drop in f found in the experiments.
According to the simulations, \(\bar_\text \), \(\bar_\text \) and \(\bar_\text \) are unlikely candidates since they had close to no effect on the \(f-I\) curve. The conductances \(\bar_\text \), \(\bar_\text \) and \(\bar_\text \) and the reversal potentials \(E_\text \) and \(E_\text \) are unlikely candidates since varying them introduced large shifts in the onset of the \(f-I\) curves not observed by Tewari et al. \(f-I\) curves resembling those in Fig. 1 could be obtained by upregulating the Ca\(^\) conductances \(\bar_\text \) and \(\bar_\text \). However, such upregulations are in conflict with previous experimental studies suggesting that PNN degradation should rather lead to a down-regulation of the mechanisms in question. Ruling out the above parameters, we are left with four possible candidate mechanisms: the reversal potentials \(E_}\) and \(E_}\), and the conductances \(\bar_\text \) and \(\bar_\text \).
Upregulating the conductances \(\bar_\text \) and \(\bar_\text \), both present in the Allen PV cell models, had an effect on the \(f-I\) curve similar to those seen in Fig. 1. Among these, upregulation of \(\bar_\text \) by PNN degradation is supported by previous experiments, while we found no mention in the literature of PNN effects on \(\bar_\text \). Likewise, increasing \(E_}\) and \(E_}\) also lead to the desired reduction in f. PNNs have been shown to accumulate cationic molecules and may provide ion sorting on neuronal membranes (Morawski et al., 2015; Burket et al., 2017). The notion that PNN degradation should affect ionic reversals reversal potentials is thus not unlikely.
Fig. 6
Frequency-input curves of Allen model 1 when varying \(c_\text \) and A \(E_}\), \(\bar_}\) and \(\bar_}\), B \(E_}(t=0)\) and \(\bar_\text \), C \(E_}(t=0)\), \(\bar_\text \) and \(\bar_\text \), D \(\bar_\text \) and \(\bar_\text \), E \(E_}(t=0)\), \(E_}\), \(\bar_\text \) and \(\bar_\text \), F Relative difference between each parameter combination and default at the largest current that gave sustained firing in both cases. The horizontal dashed line indicate the relative difference between f of Sham and GBM22 in Tewari et al.’s experiments. The difference between Sham and GBM14 is a bit larger. Default - default values, \(E_\text =53\) mV and \(E_\text (0)=131.06\) mV. For the altered models, \(E_\text =63\) mV and \(E_\text (0)=161.53\) mV, \(c_\text \) is increased by a factor 1.5 and the conductances are indicated in the legend
As shown in Fig. 6, the experiments of Tewari et al. could be explained through various combinations of changes in a selection of the parameters \(c_\text \), \(_}\), \(_}\) \(\bar_}\) and \(\bar_\text \). Allen model 1 was chosen as that yielded the strongest responses to changes in parameters, and was therefore the most promising candidate for recreating the 38% average drop in f from Tewari et al.’s experiments.
In general, achieving a reduction in f similar to what was seen in Fig. 1 required quite large changes in several parameters, and a large increase in \(c_\text \) was a necessary part of it. In Fig. 6, \(c_\text \) was increased by a factor 1.5, \(E_}\) and \(E_}\) (when included) were shifted by 30 and 10 mV, respectively, while \(\bar_}\) and \(\bar_\text \) were varied (jointly, when both were included) by factors between 1.5 and 4 as indicated in the figure legends. Upregulation of \(\bar_\text \) by such a high factor due to PNN degradation is supported by the experiments by Dembitskaya et al. (2021). It was there found that on average, \(\bar_\text \) increased by a factor three after PNN degradation, but changes up to a factor six was within the standard deviation in the experimental data. For the remaining parameters, the literature gives no guidance as to whether PNN degradation should affect them in the way suggested in Fig. 6.
Not surprisingly, the largest effect on f was found when the full set of candidate mechanisms were changed in the same model. When \(\bar_}\) and \(\bar_\text \) were increased by a factor four (relative to their default values in the model), the reduction in f exceeded that seen in Fig. 1.
留言 (0)