
{kind=link}
記住我
The immunosuppressive tumor microenvironment (TME) represents one of the main reasons for the poor outcome of immunotherapy against cancer. Hence, exploring efficient methods to activate immune cells in immunosuppressive environment is a crucial task for the development of better immunotherapies. Dendritic cells (DCs) are a heterogeneous group of specialized antigen-presenting cells that play a vital role in initiating and regulating innate and adaptive immune responses.1 In the TME, DCs acquire, process, and present tumor-associated antigens on MHC (major histocompatibility complex) molecules, and provide co-stimulation and soluble factors to elicit T cell responses.2 In clinical settings, many therapy strategies have been developed to target DCs, such as delivering antigens, mobilizing and activating endogenous DCs using stimulant adjuvants, and generating DC-based vaccines.3 4 Nevertheless, to achieve more effective antitumor immunotherapy, strategies to accurately target DC and avoid immune tolerance of DC in the TME are needed.
In recent years, considerable attention has been attracted by the critical roles of mitochondria within the TME, including metabolism-regulation and immune-activation. On the one hand, mitochondria participate in the regulations of cellular metabolism, which produces reactive oxygen species (ROS). It has been well documented that ROS is an important factor affecting the proliferation, metastasis, and stemness of tumor cells.5 On the other hand, mitochondria could activate immune response through damage-associated molecular patterns (DAMPs). According to ‘endosymbiont hypothesis’, it is very likely that mitochondria are evolved from aerobic prokaryotes.6 They possess many features of their bacterial ancestors, including a circular genome containing CpG DNA and the ability to form N-formyl peptides.7 It has been reported that mitochondrial components could efflux into the cytoplasm or extracellular space in specific conditions to be recognized as DAMPs by the corresponding pattern recognition receptors.8 During the process, circular genome containing CpG DNA plays key role in the activation of immune response. Compelling evidence suggests that the release of mitochondrial DNA (mtDNA) from necrotic cells activates the NF-κB signaling pathway and stimulates expression of other pro-inflammatory genes by interacting with the neutrophil TLR9 receptor, resulting in a corresponding inflammatory response in vitro.9 Besides, our previous work has demonstrated that the oxidized mtDNA from tumor cells could activate STING signaling and then induce the tumor specific immune response.10
Most mitochondrial proteins are encoded by nuclear DNA, translated in the cytoplasm, and then transported to the corresponding functional sites in the mitochondria.11 The mitochondrial transcription factor A (TFAM) is the most abundant protein associated with mtDNA encoded by nuclear genes.12 This protein not only initiates mtDNA transcription and replication but also maintains mtDNA structure.13 Interestingly, the role of TFAM in mitochondria is similar to the role of histones in nucleosome. TFAM wraps mtDNA entirely to form a nucleoid structure14 that may protect mtDNA against ROS.15 Tissue-specific ablation of TFAM has been used to mimic the mitochondrial dysfunction observed in various human diseases.16 17 Previous studies on TFAM in tumor models mainly focused on how it affects metabolism, proliferation, and invasion of tumor cells.18 However, a study to reveal how TFAM in immune cells contributes to remodeling of the TME is absent.
In the current study, we used the myeloid-specific Tfam knockout mice to investigate how TFAM contributes to antitumor immunity and the underlying mechanism. Interestingly, Tfam deficiency in myeloid cells led to preferable activation of DCs. The enhanced antitumor humoral and cellular immunity were provoked by Tfam deficient DCs through cGAS-STING pathway elicited by mtDNA leakage. Additionally, the tumor growth and immune microenvironment were characterized in myeloid-specific Tfam knockout mice. This study may indicate the critical role of TFAM in balancing immunity and metabolism in DCs, which provides more insights for tumor immunotherapy targeting mitochondria.
MethodsAnimalsSix-to-eight-week-old C57BL/6 mice were purchased from Beijing Vital River Laboratory Animal Technology Company, and C57BL/6 mice with Tfam-floxed alleles, OT-I, or expressing Cre-lyz2 were purchased from Jackson Lab (Stock#026123, stock#003831 and Stock#004781, respectively). The Tfamflox/flox mice were crossed with transgenic mice expressing Cre-lyz2 as lyz2-Cre/Tfamflox/flox or Tfam-/-. Tfamfl/fl littermates not expressing Cre and wild type (WT) were used as control mice in this study. The mice were housed in a specific pathogen-free environment.
Statistical analysis and reproducibilityAll of the experiments (including the western blots) were repeated independently at least twice with similar results. All data were analyzed using one-way analysis of variance (ANOVA), two-way ANOVA or two-sided Student’s t-test (GraphPad InStat Software, California, USA). Log-rank (Mantel-Cox) test was used for survival curves analyses. Results were presented as the means±SEM. The statistic differences were denoted as *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, and p<0.05 was considered significant.
Other details and additional experimental procedures are provided in online supplemental methods.
Results
Deletion of Tfam in myeloid lineage inhibits lung tumor progressionTo investigate the role of TFAM in immune cells of cancer, we deleted TFAM in the myeloid cell lineage by crossbreeding Lyz2-Cre and Tfamfl/fl mice (online supplemental figure 1A,B), referred to as Tfam-/-. Tfamfl/fl littermates not expressing Lyz2-Cre and WT were used as control for the study. Murine pulmonary metastasis models were established by injecting LLC (Lewis lung carcinoma) or B16-F10 cells intravenously. Interestingly, we found that tumor growth and metastasis were significantly reduced in myeloid cell-specific TFAM knockout groups in both models (figure 1A–C; online supplemental figure 1C). Inhibited tumor progression was detected by measuring the lung weight, metastatic area, and metastatic nodules, which resulted in the prolonged survival of Tfam-/- tumor-bearing mice (figure 1C). Previous studies have claimed that the tumor could grow up to a diameter of 2–3 mm without their tumor microvascular system,19 and the deletion of Tfam in myeloid cells significantly reduced the metastatic nodules with diameter >3 mm (online supplemental figure 1D). Moreover, representative immunohistochemical staining of cleaved caspase-3 and CD31 were analyzed to observe tumor cell apoptosis and tumor angiogenesis. The Tfam-/- group showed higher expression of cleaved caspase-3 and lower expression of CD31 in the tumor sections (figure 1D,E; online supplemental figure 1E,F), suggesting more apoptosis tumor cells and fewer numbers of CD31+ tumor vessels compared with the control group. These results revealed that knockout of TFAM in myeloid lineage effectively inhibited tumor growth and prolonged survival in mice.
 Figure 1
Figure 1 Low expression of Tfam in myeloid inhibits lung tumor growth. (A, B) Tfam deletion in myeloid inhibited tumor growth in LLC (A) or B16-F10 (B) lung metastatic tumor models. LLC cells (5×105) or B16-F10 cells (2×105) were intravenously injected into control or Tfam-/- mice to establish experimental pulmonary metastasis models (n=6–9 mice), Tfamfl/fl littermates and wild type were used as control. Mice were sacrificed on day 24 (LLC models) or day 14 (B16-F10 models), and pulmonary physiology was evaluated, including gross images and H&E staining of lung, measurement of metastatic area (n=3 mice’s lungs were paraffin embedded) and nodules (n=6–9 mice). Scale bars represent 2 mm. (C) Tfam deletion in myeloid prolonged the survival of tumor-bearing mice. Survival statistics of mice from LLC (5×105) or B16-F10 (5×105) lung metastatic tumor models (n=10–11 mice). (D, E) Immunohistochemical staining of cleaved caspase-3 (D) or CD31 (E) in lungs of the mice described in (A, B). Scale bars represent 20 µm. Data are represented as mean±SEM. Statistical significance in (A, B) was determined by a two-sided unpaired t-test. Survival curve data in (C) were analyzed by log-rank (Mantel-Cox test). Representative results in (A–C) and pictures in (A, B, D, E) from two independent experiments are shown. *p<0.05, ***p<0.001. LLC, Lewis lung carcinoma.
Tfam deletion activates tumor immune microenvironment with increased lymphocyte infiltrationTo further study the effects of Tfam deletion on tumor growth, we next investigated whether and how TFAM deficiency in myeloid lineage would modulate tumor immune microenvironment (TIME). We used flow cytometry (FCM) to analyze the lung tissues from tumor-bearing mice inoculated with LLC cells. DCs are known to critically influence the adaptive immune response against tumors. In the current study, the percentages of DCs in Tfam-/- mice were significantly increased when compared with control group (figure 2A). Interestingly, the infiltration of alveolar macrophages and tumor-associated macrophages (TAMs) was not altered (figure 2B,C). Furthermore, TFAM deficiency didn’t obviously influence the polarization of TAMs (figure 2C). In the meantime, tumor-infiltrating monocytes, as well as Ly6Cmid monocytes, and neutrophils were assessed, and the increased population of the infiltrating innate cells suggested an activated inflammatory immune response (online supplemental figure 2A).20 Furthermore, the efficiency of TFAM knockout in DCs from the TME was also confirmed by FCM, and the TFAM expressions in DCs were dramatically reduced in the lungs of Tfam-/- mice (online supplemental figure 2B).
 Figure 2
Figure 2 TFAM deletion in myeloid transforms tumor immune microenvironment and increases lymphocyte infiltration. (A–F) LLC cells (5×105) were intravenously injected into control or Tfam-/- mice to establish experimental pulmonary metastasis models. Mice were sacrificed on day 24 to collect the lungs. Then the single-cell suspension of the whole lungs with metastatic tumors was prepared and subjected to FCM analysis. (A) Representative scatterplots of the gated DCs (CD45+ CD3- CD11b+ CD11c+ MHCII+ CD24+) are shown in the left panel and quantified in the right panel (n = 4 mice). (B) The percentages of alveolar macrophages (CD45+ CD3- CD11b- CD11c+) are quantified (n = 4 mice). (C) The percentages of TAMs (CD45+ CD3- CD11b+ F4/80+), M1 TAMs (CD45+ CD3- CD11b+ F4/80+ MHCII+), and M2 TAMs (CD45+ CD3- CD11b+ F4/80+ CD206+) are quantified (n = 4 mice). (D) The percentages of CD3+ CD8+ CTLs gated from CD45+ cells (left panel) and activated CD69+ CTLs gated from CD45+ CD3+ CD8+ (right panel) are quantified (n=5 mice). (E) The percentages of GzmB+ or IFN-γ+ T cells are quantified. Cells are gated from CD45+ CD3+ CD8+ subpopulation (n=5 mice). (F) The percentages of CD25+ or PD-1+ T cells are quantified. Cells are gated from CD45+ CD3+ CD4+ subpopulation (n=5 mice). (G, H) Immunofluorescence staining of CD45 (green), CD3 (red), CD8 (green) and DAPI (blue) in lungs of the mice from pulmonary metastasis models of LLC (G) or B16-F10 (H). Scale bars represent 20 µm. (I) B16-F10 cells (2×105) were intravenously injected into control or Tfam-/- mice to establish experimental pulmonary metastasis models. Mice were sacrificed on day 14, then the single cell suspension of the whole lung with metastatic tumor was prepared and subjected to FCM analysis. Representative scatterplots of the p15E-specific CD8+ T cells are shown in the left panel and quantified in the right panel. Cells are gated from CD45+ CD3+ subpopulation (n=4 mice). Data are presented as mean±SEM. Statistical significance in (A–F, I) was determined by a two-sided unpaired t-test. Representative results in (A–F, I) and pictures in (G, H) from two independent experiments are shown. *p<0.05, ***p<0.001, NS, not significant. LLC, Lewis lung carcinoma. FCM, flow cytometry; DC, dendritic cell; MHC, major histocompatibility complex; TAMs, tumor-associated macrophages.
Besides immune myeloid cells, the populations of T cells play dominant roles in activating antitumor immunity. We assessed how TFAM knockout in myeloid cells affects the tumor-infiltrating T cells in LLC lung metastasis models. As shown in figure 2D, the elevation in the percentages of both CD8+ cytotoxic T lymphocyte (CTL) and activated CTLs (CD8+ CD69+ T cells) were observed. Furthermore, Tfam-/- mice also exhibited a significant increase in the percentages of CD8+ IFN-γ+ T cells and CD8+ GzmB+ T cells (figure 2E). Elevated expression of IFN-γ and granzyme B (GzmB) in activated T cells are generally considered as the favorable condition to provoke a more effective antitumor immunity.21 As for T helper cells, CD4+ CD25+ and CD4+ PD-1+ T cells were significantly diminished in Tfam-/- mice when compared with that of the control group as detected (figure 2F). Moreover, immunofluorescence staining of lung sections also confirmed the significant increase of tumor-infiltrating CD45+, CD3+, CD8+ lymphocyte cells in LLC or B16-F10 lung metastasis models (figure 2G,H), which did not undergo apoptosis (online supplemental figure 2C,D). We further investigated whether the CD8+ T cells could specifically recognize p15E, an endogenously expressed antigen in B16-F10. As shown in figure 2I, we found that the induction of p15E-specific CD8+ T cells was significantly increased in Tfam-/- group in B16-F10 lung metastasis tumor models (figure 2I). These results demonstrated that TFAM knockout in myeloid cells might significantly enhance immune cell infiltration and T cell activation, which might result in a reshaped TME for boosting host immune responses to kill tumor cells.
TFAM deficiency activates DCs in vitro and in vivoTo investigate how Tfam deficiency in myeloid linage affects the cell characteristics and how they stimulate TME, we further investigated the effect of TFAM deletion in a population of myeloid-derived cells, especially DCs, which are responsible for the antigen presentation and antitumor immunity activation. It is known that DCs express several surface receptor costimulatory molecules after the exposure to antigens or inflammatory stimulants, which induces the maturation, migration of DCs and enhances the antigen-presenting process to primary T cells in secondary lymphatic organs.22 To simulate the tumor condition in vitro, we used a conditioned medium containing tumor supernatant (TS) of LLC cells to stimulate primary mouse bone marrow-derived DCs (BMDCs), whose phenotypes and TFAM recombination efficiency were analyzed by FCM (online supplemental figure 3A–D). The identified studies indicate that the TS could mediate DC activation.23We found that the expression of costimulatory molecules on DCs including CD40, CD86, and MHCII were significantly elevated in the Tfam-/- group when compared with the control (figure 3A). This process partially relied on nucleic acid, because SuperNuclease repressed the maturation of DCs induced by TS stimulation (online supplemental figure 4A–C). Moreover, cytokine levels such as TNF-α, IL-6, IL-1β, and IL-12 p40 were also heightened in the Tfam-/- mice derived BMDCs after TS stimulation (figure 3B). In addition, TFAM deletion also notably up-regulated the phagocytosis capacity of the BMDCs after TS stimulation as assessed by the uptake of FITC-dextran (figure 3C,D).
 Figure 3
Figure 3 TFAM deficiency activates DC both in vitro and in vivo. (A) Tfam deficiency promotes the maturation of DCs before or after TS stimulation in vitro. BMDCs from control or Tfam-/- mice were stimulated with or without LLC tumor supernatant (TS) for 24 hours, then subjected to flow cytometry analysis to detect the expression of costimulatory molecules. The upper panel shows the representative histograms of the gated CD11c+ DCs (n = 3 biologically independent samples). The quantitative data of flow cytometry results are shown in the lower panel. (B) Tfam deficiency promotes the secretion of inflammatory cytokines of DCs after TS stimulation for 24 hours in vitro. Levels of TNF-α, IL-6, IL-1β, and IL-12 p40 in the supernatant from BMDCs treated as in (A) were detected by ELISA (n = 3 biologically independent samples). The original levels of cytokines in TS were subtracted. (C) Tfam deficiency promotes the antigen uptake of DCs in vitro. BMDCs were stimulated with or without TS for 24 hours, followed by incubation with 1 mg/mL FITC‐Dextran for 1 hour at 37°C and then analyzed under a fluorescent microscope. Scale bars represent 10 µm. (D) The mean fluorescence intensity (MFI) of CD11c+ BMDCs in (C) was further analyzed by flow cytometry (n = 3 biologically independent samples). The control group was performed by co-culturing BMDCs with FITC-dextran at 4°C. (E) Tfam deficiency promotes the antigen presentation of DCs in vitro. Representative scatterplots of the gated CD8+ T cells from OT-I mice are shown in the left panel and quantified in the right panel. Numbers indicate the percentage of proliferated CFSE-negative CD8+ T cells from OT-I mice (n = 3 biologically independent samples). (F) Tfam deficiency promotes the migration and maturation of DCs in vivo. Single-cell suspension of lymph nodes from control or Tfam-/- mice were subjected to flow cytometry analysis. The percentages of CD197+, MHC II+, CD40+, or CD80+ of CD11c+ DCs are quantified from CD45+ gated subpopulation (n=6 mice). Data are presented as mean±SEM. Statistical significance was determined by two-way ANOVA in (A, B, F) or a two-sided unpaired t-test in (D, E). Representative results in (A, B, D, F) and pictures in (C) from three independent experiments are shown. Representative results in (E) from two independent experiments are shown. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ANOVA, analysis of variance; BMDCs, bone marrow-derived dendritic cells; MHC, major histocompatibility complex. CFSE, carboxyfluorescein succinimidyl ester.
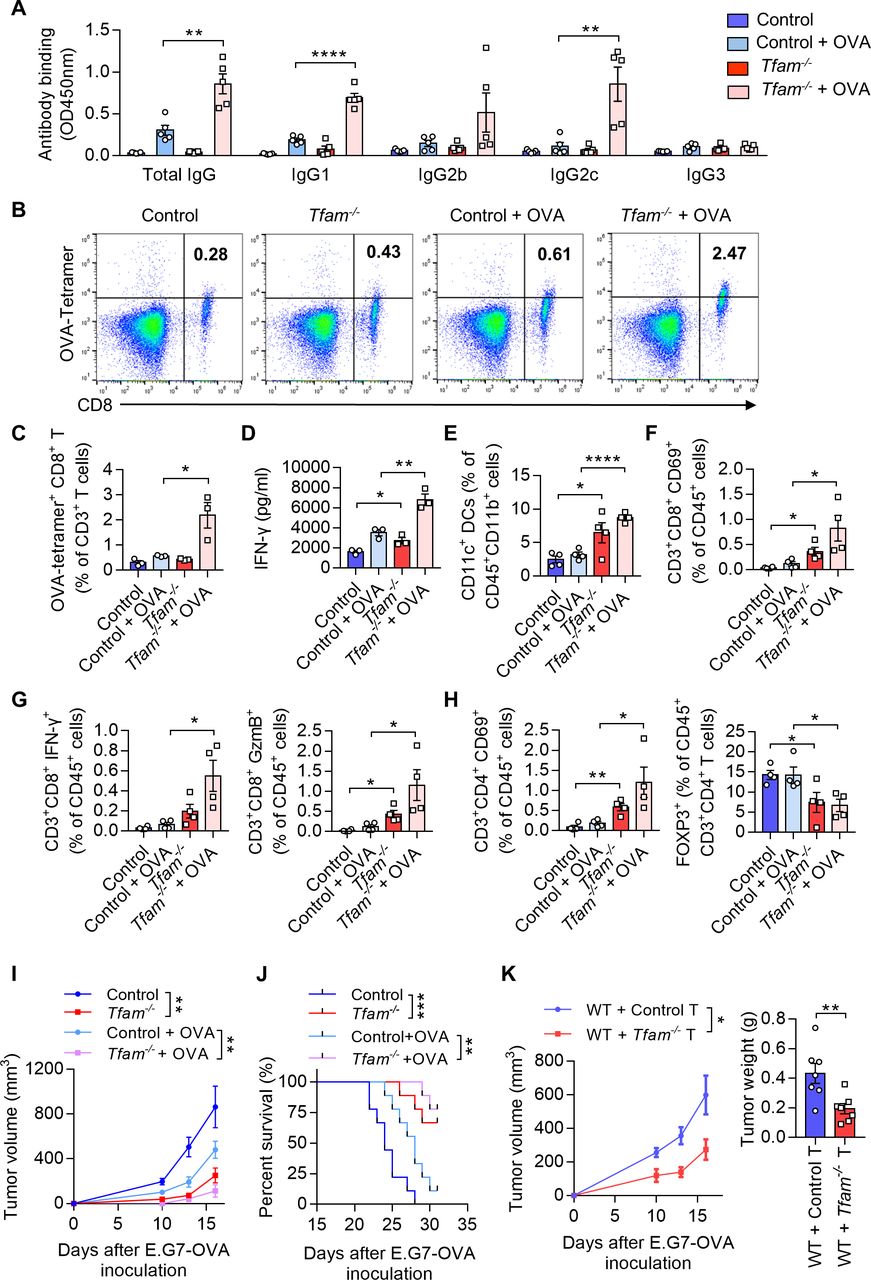 Figure 4
Figure 4 TFAM deficient DC enhanced specific humoral and cellular immune responses. (A) Tfam deficiency enhances the specific anti-OVA humoral immunity. Control and Tfam-/- mice were vaccinated subcutaneously three times with or without 10 µg OVA antigen in PBS on days 0, 14, and 21. The mouse serum were collected on day 28 and levels of the total IgG and IgG subclasses were determined by ELISA. Serum antibody binding was determined by absorbance at 450 nm (n=5 mice). (B–D) Tfam deficiency enhances the specific anti-OVA cellular immunity. Splenic lymphocytes from immunized mice in (A) were isolated on day 28 and further incubated in vitro with CD8+ specific OVA257–264 peptides (10 µg/mL) for 72 hours. The generation of CD8+ CTLs was determined by FCM using PE-conjugated H-2Kb/OVA257–264 tetramer. Representative scatterplots of the OVA-specific CD8+ T cells gated from CD3+ cells are shown in (B) and quantified in (C). Level of IFN-γ in the supernatant was measured by ELISA (D) (n=3 mice). (E) Frequency of CD11c+ DCs in the spleen gated from CD45+ CD3- CD11b+ are determined (n=4 mice). (F) Frequency of CD3+ CD8+ CD69+ CTLs in the spleen gated from CD45+ are determined (n=4 mice). (G) Frequency of CD3+ CD8+ IFN-γ+ CTLs in the spleen gated from CD45+ are determined in the left panel, and frequency of CD3+ CD8+ GzmB+ CTLs in the spleen gated from CD45+ are determined in the right panel (n=4 mice). (H) Frequency of CD3+ CD4+ CD69+ T cells in the spleen gated from CD45+ are determined in the left panel, and frequency of FOXP3+ T cells in the spleen gated from CD45+ CD3+ CD4+ are determined in the right panel (n=4 mice). (I) Tfam deficient potentiates the antitumor effect of OVA vaccine in vivo. In the prophylactic model, control or Tfam-/- mice were immunized as in (A) (n=5 mice) and then injected subcutaneously with E.G7-OVA cells (5×105) 1 week after the third immunization. Tumor growth was monitored at the indicated times. (J) Tfam deficiency potentiates the survival of OVA-vaccinated tumor-bearing mice. Control and Tfam-/- mice were immunized with OVA and injected with E.G7-OVA (5×105) as previously described. Survival of mice was monitored daily (n=9 mice). (K) In the cellular adoptive therapy model, CD8+ T lymphocytes were isolated from immunized mice as in (A) on day 28 and subsequently injected intravenously into recipient mice, which were wild type mice subcutaneously inoculated with E.G7-OVA (5×105) cells. Tumor growth was monitored at the indicated times (left panel) and tumor weight was recorded after sacrifice on day 16 post-transplantation (right panel) (n=7 mice). Data represent the mean±SEM. Statistical significance was determined by two-way ANOVA in (I), left panel of (K) or a two-sided unpaired t-test in (A, C–H, right panel of K). Survival curve data in (J) were analyzed by log-rank (Mantel-Cox test). Representative results in (A–K) from two independent experiments are shown. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ANOVA, analysis of variance; DC, dendritic cell; i.v., intravenous.
To analyze the ability of presentation in BMDCs, primary BMDCs from mice immunized with OVA were co-cultured with CFSE (Carboxyfluorescein succinimidyl ester) -labeled CD8+ T lymphocytes from OT-I mice, whose TCR can specifically recognize OVA via MHCI molecule. CFSE is a fluorescent dye that is equally partitioned during cell division. FCM analysis showed an elevated rate of CFSE which indicated TFAM deficiency in BMDCs could significantly contribute to the antigen presentation and the proliferation of specific CD8+ T cells (figure 3E). In the next set of experiment, we examined the migration and activation of DCs in the lymph nodes in Tfam-/- and control mice in vivo. The chemokine receptor CCR7 (CD197) connects innate and adaptive immunity and plays a vital role in the homing of DCs to secondary lymphatic organs, enabling rapid proliferation and differentiation of T cells.24 The results suggested an elevated expression of CD197 in DCs harvested from the lymph nodes of Tfam-/- mice when compared with that of the control groups by FCM (figure 3F). In addition, the lymphoid DCs also demonstrated the increased levels in CD40, CD80, and MHCII expression in Tfam-/- group, which indicated that deletion of TFAM could effectively promote the maturation and migration of DCs in vivo (figure 3F). These results suggest that the TFAM deletion in DCs leads to enhanced maturation and activation of DCs, promoting the priming process and the proliferation of T cells.
TFAM deficient DCs enhance humoral and cellular immune responses in vivoAs TFAM deletion resulted in enhanced maturation and activation of the DCs in vitro, we next investigated whether the humoral and cellular immune responses were promoted in Tfam-/- mice. To better model tumor responses, OVA was used as the model antigen to validate the activation of immune responses in vivo. Tfam-/- and control mice were immunized with OVA for three times and the sera were collected. The levels of total IgG and its subclasses were measured. The results showed that the antibody titers of IgG, IgG1, and IgG2c were all elevated in immunized Tfam-/- mice when compared with that of the control group (figure 4A), which suggested a potent activation of humoral immunity in mice Tfam-/- BMDCs. When it comes to the cellular immune response, we demonstrated that Tfam-/- deletion simulated more effective OVA-specific CD8+ T cell responses as evidenced by the increase in spleen lymphocytes from Tfam-/- and control mice after the stimulation with OVA257-264 peptides (figure 4B,C). In accordance, the ELISA results showed the increased production of IFN-γ by CD8+ T lymphocytes in Tfam-/- cells after OVA immunization when compared with the control (figure 4D). Furthermore, the immune-microenvironment and the critical cell populations in the spleen of Tfam-/- and control mice were characterized by FCM. The increased populations of DCs (CD11c+ DCs, figure 4E), CD8+ CD69+ T cells (figure 4F), CD8+ IFN-γ+ T cells, CD8+ GzmB+ T cells (figure 4G), CD4+ CD69+ T cells (figure 4H) and the decreased percentages of CD4+ FOXP3+ cells were also detected (figure 4H).
Next, we used the prophylactic model to identify the anti-cancer immunity activation in Tfam-/- and control group. Mice were challenged with subcutaneous injection of E.G7-OVA cells after three times of immunization. The tumor growth was significantly inhibited in Tfam-/- mice in the prophylactic tumor model with the OVA immunization when compared with that of the control mice (figure 4I), and this group of mice showed prolonged survival (figure 4J). To investigate whether such tumor inhibition effect was due to the activation of cellular immune response, the adoptive transfer study was carried out by isolating CD8+ T lymphocytes from the immunized mouse spleen. CD8+ T lymphocytes from immunized Tfam-/- mice or control mice were injected intravenously in WT mice on day 1 before E.G7-OVA cell inoculation and on day 1 and day 3 after E.G7-OVA cell inoculation. Interestingly, the tumor growth as detected by tumor weight and tumor volume was significantly inhibited in the group that received the adoptive transfer CD8+ T cells from Tfam-/- mice when compared with that received the control CD8+ T cells (figure 4K). To further figure out the myeloid cell subset responsible for the activation of antitumor immunity, we adopted control or Tfam-/- DCs/macrophages to WT LLC tumor-bearing mice. As expected, adoption of Tfam-/- DCs instead of Tfam-/- macrophages significantly inhibited tumor metastasis and tumor growth (online supplemental figure 5). In summary, TFAM deficiency not only caused DC activation, but also led to more efficient activation of antitumor humoral and cellular immunity in vivo.
留言 (0)