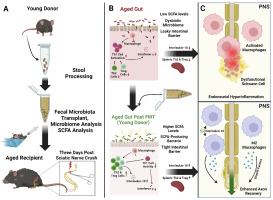
The structure of the peripheral nervous system (PNS) is extremely complex due to the myriad connections it establishes with the central nervous system (CNS) and with tissues and organs of the thoracic and abdominal cavity of the body. The PNS consists of two components: the afferent sensory system, which is responsible for conveying information to the CNS from the body itself and the external environment, and the efferent motor system, which carries information from the CNS to the skeletal muscles. In the PNS, the nerves contain the axons of the sympathetic ganglions, motor neurons and dorsal root ganglia or DRG, which define the identity of the sensory stimulus. Unique to DRG neurons, both branches departing from DRG cell bodies are considered axons and the sensory stimuli lead to the generation of action potentials in the terminal of the peripheral branch that are conducted toward the stem axon bifurcation (Fig. 1). From this point, action potentials are transmitted along the central axon towards the CNS. In this way, the peripheral DRG axon resembles a dendrite, as the action potential is generated at the peripheral terminal and propagates towards the bifurcation of the stem process. In contrast, the central process is more classically axon-like, as it conducts the signal from the bifurcation to the CNS.
DRG axons that are wrapped by myelin lamellae provided by Schwann cells can extend from a few millimeters to up to one meter in humans, for example, from the spinal cord to the toes, and are highly dependent on energy metabolism (Corfas et al., 2004). This energy dependency requires a well-constructed machinery to shuttle vital cellular components from the cell body along the axonal projections and vice versa. Axons of peripheral nerves also contain non-myelinated Remark bundles that wrap small caliber axon (Jessen and Mirsky, 2005). Many of them are sensory c-fibers that transmit pain information to the brain. The alteration of these fibers, even in the absence of injury, is associated with hypersensitivity to mechanical and thermal stimuli and in the alteration of motor coordination (Faroni et al., 2014; Harty and Monk, 2017).
Unlike the CNS, which is protected by bone, meninges, and cerebral fluid, the PNS has no blood barrier to protect itself from toxins and pathogens in the blood. Therefore, due to its long reaching and complex structure, the PNS is exposed to many environmental factors, genetic predispositions, and metabolic diseases, including diabetes. The PNS can also be the target of different viruses such as herpes zoster, rabies virus and human immunodeficiency virus (HIV) (Koyuncu et al., 2013). Lastly, the PNS can be poisoned by toxins and drugs, including chemotherapeutic compounds.
Any disorder affecting the PNS, defined as peripheral neuropathy (PN), is highly debilitating because it disrupts movement and impairs peripheral sensitivity. PNs are typically classified by the localization of the nerve injury, the type of fibers involved (small or larger diameter; myelinated or unmyelinated) and the nature of the neuropathic trigger as summarized in Table 1 (Alberti et al., 2019; Bansal et al., 2006; Beadon and Léger, 2020; Benson and Kincaid, 2007; Chin and Latov, 2005; Collins and Hadden, 2017; De Jonghe et al., 1997; Duyff et al., 2000; Ebenezer et al., 2007; Gainsborough et al., 1991; Ho et al., 1998; Karam and Dyck, 2015; Köller et al., 2005; Lewis and Howdle, 2003; Meier and Moll, 1982; Nobile-Orazio, 2013; Rotthier et al., 2012; Said, 2013; Staff et al., 2017; Takada et al., 2012; Weis et al., 2017; Zambelis et al., 2005).
Despite this clinical knowledge, the mechanisms underlying the etiology of most PNs are poorly understood, creating a burden for the development of neuroprotective therapies. Among several candidates, neuronal microtubules (MTs) remain one of the most likely culprits for several reasons. First, loss of the stable MT pool is thought to participate in neurodegenerative and neuropathic disease through its well-established role in long distance cargo transport and in providing structural support. Second, recent evidence suggests that dynamic and unmodified MTs can modulate signaling events, synaptic plasticity and neurotransmission, and that perturbation of MT dynamic behavior may directly contribute to neurodegeneration (Parato and Bartolini, 2021; Peris et al., 2022; Waites et al., 2021).
An increasing number of functional studies have underscored the physiological roles of tubulin post-translational modifications (PTMs) that accumulate on stable MTs in different cells and tissues, including neurons. MTs change their properties when they accumulate tubulin PTMs, including regulation of axonal transport, indicating that unbuffered fluctuations in tubulin PTMs can drive neurological disease. Interestingly, paclitaxel-induced MT hyperstabilization was recently found to sever the connection between sensory neurons and the extracellular matrix, providing evidence that regulation of tubulin PTMs may also guide proper localization of the machinery that allows neurons to interact with the extracellular matrix via integrin receptors (Shin et al., 2021).
A strong link between damage to the MT cytoskeletal and neurodegeneration of the axon is supported by multiple observations. The MT cytoskeleton is crucial in maintaining the asymmetric shape and polarity of DRG neurons, which have an important central-peripheral polarity regarding electrophysiological properties. Indeed, MTs promote transmission of sensory information, which travels through the peripheral axon of the nerves to the skin, where free sensory endings can emerge and branch into an arbor that receives and integrates information.
Furthermore, MTs have a direct impact on the normal functioning of axonal transport by affecting motor proteins and the distribution of mitochondria, the organelles responsible for ATP generation, Ca2+ buffering and phospholipid synthesis at contacts with the endoplasmic reticulum (ER). In particular, mitochondria homeostasis is regulated by a process of mitochondrial fusion and fission, transport, and heterotypic contacts with other organelles. This series of steps has been termed “mitochondrial dynamics” and is fundamental for regulating axonal viability (Detmer and Chan, 2007). Both tubulin and MTs have been implicated in the regulation of all aspects of mitochondrial dynamics, indicating a strong interplay between tubulin damage and failure of mitochondrial homeostasis.
In this Review, we summarize what we currently know about the physiological role of tubulin PTMs in the PNS in healthy conditions and the implication of selected tubulin PTMs in the etiology of both familial and toxic forms of peripheral neuropathy.
MTs are elements of the neuronal cytoskeleton with fundamental roles in the maintenance of axonal integrity and polarity. Peculiar to their intrinsic nature, MTs can adjust their own structure and organization in response to the cell’s physiological needs. At an ultrastructural level, MTs are polarized polymers derived from the regulated addition of α/β tubulin heterodimers into 13 protofilaments that arrange around a hollow core (Fig. 2). Both α and β tubulins exist in numerous isotypic forms encoded by different genes, contributing to the plasticity of MTs in adapting to specific cellular functions or physiological demands. Dynamic MTs are characterized by a fast growing plus end with β-tubulin exposed and a slow growing minus end with α-tubulin exposed. After polymerization, GTP is hydrolyzed and the affinity of tubulin for adjacent molecules decreases, promoting depolymerization. The frequency at which MT polymerization switches to depolymerization and vice versa, and the speed of MT growth and shrinkage, provide the basic criteria to describe intrinsic dynamic MT behavior, also known as “dynamic instability” (Mitchison and Kirschner, 1984).
Neurons possess both dynamic and stable MTs, and both their relative frequency and density along the neurite are functionally important, with stable MTs remaining in a relatively constant polymerized form (Kapitein and Hoogenraad, 2015; Katrukha et al., 2021). Axons contain bundles of long stable MTs which act as roads and provide support for the long-distance transport of proteins. Besides the intrinsic properties of tubulin, dynamic instability is regulated by numerous MT-associated proteins (MAPs), MT-severing proteins, MT plus-end tracking proteins (+TIPs), and motor proteins such as kinesin-1 or dynein.
In addition to dynamic instability, MT polarity is critical to support neuronal functions which rely on the early establishment of non-centrosomal MT arrays. Any change of MT polarity may influence the geometry of the neuron and affect the orientation of organelles and cargos. This is because MT polarity is recognized and regulated in part by the molecular motors dynein and kinesin that can only move in one direction and play a role in constructing the tracks that they use to carry cargos (Kelliher et al., 2019; Meiring et al., 2020; Rao et al., 2017; Vale, 2003).
The polarity of MTs is established during neuronal development. For example, in rat cortical neurons, initially, about 80% of axonal MTs point toward the growth cone, but in mature axons, almost all MTs do so (Baas et al., 1989; Baas et al., 1988; Yau et al., 2016). In vertebrates, neuronal MTs themselves are transported unidirectionally in the axon with the plus ends directed away from the cell body and bidirectionally in dendrites where they form a mixed polarity of parallel and antiparallel arrangements (Baas et al., 2006; Rolls and Jegla, 2015; Thyagarajan et al., 2022). The adjustment of polarity is also observed in association with neurite extension, retraction, growth cone progression and in axonal injury (Banker, 2018; Witte et al., 2008). Drosophila neurons, for example, can readjust their polarity after axonal injury by converting a dendrite to a new axon. This process, known as the “re-specification of axons”, is preceded by an increase in the polymerization of MTs, which is a fundamental process of compensation and/or adaptability in the mechanism of regeneration (Stone et al., 2010).
In neurons, MTs are the main component of the axon initial segment (AIS), a highly specialized neuronal compartment in the proximal axon with key functions in the initiation of action potentials and maintenance of neuronal polarity by restricting the clustering of Na+ channels, membrane proteins and cell adhesion molecules. In DRG neurons, the AIS resides in the proximal unmyelinated region and contains a higher density of loosely packed MTs connected by lateral cross bridges (Nascimento et al., 2018). The difference in MT density between the proximal and distal axon is believed to play a role in the movement of cargos (Farías et al., 2015; Harterink et al., 2019; Palay et al., 1968; Peters et al., 1968).






留言 (0)