
記住我



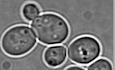

Mouse embryonic stem cells (ESCs) are derived from the inner cell mass of the pre-implantation blastocyst. These cells exhibit unlimited self-renewal capacity and, under appropriate stimuli, retain the potential to differentiate into the three germ layers (Bradley et al, 1984). Mouse ESCs are a useful model to study early mammalian development as their differentiation potential is more robust than that of ESC-like cells derived from other mammals such as humans, which exhibit primed pluripotency and represent a more advanced embryonic stage (Ginis et al, 2004; Nichols & Smith, 2009).
Zinc finger-containing proteins (ZFN or ZFP for human or mouse, respectively) are among the largest family of proteins, commonly containing a minimum of one zinc-finger (ZnF) domain, which recognizes DNA sequences with high affinity. This family of transcription factors plays important roles in a variety of cellular processes including development, cellular differentiation, metabolism and oncogenesis (Cassandri et al, 2017). Although ZNF/ZFPs were initially classified as transcription factors, several studies have highlighted additional functions of ZNFs. For instance, it has been shown that ZFP217 could recruit the methyltransferase-like 3 (METTL3) into an inactive complex and hence restrict N6-methyladenosine (m6A) deposition on pluripotency transcripts (Aguilo et al, 2015; Lee et al, 2016). In addition, recent studies identified that ZnF domains can bind RNA, and many ZNF/ZFPs act as putative RNA-binding proteins (RBPs) (Brannan et al, 2016). Indeed, analysis of quantitative global mRNA–protein interaction approaches identified ZNF207 (also termed BuGZ; Bub3 interacting GLEBS and Zinc finger domain-containing protein) as an RBP, among other ZNFs (Baltz et al, 2012; Castello et al, 2012). ZNF207 is conserved in eukaryotes. It associates with Bub3 and with spindle microtubules to regulate chromosome alignment (Jiang et al, 2014, 2015; Toledo et al, 2014). Furthermore, both ZNF207 and Bub3 interact with the spliceosome and are required for interphase RNA splicing (Wan et al, 2015), yet its specific molecular role remains elusive.
In human ESCs, ZNF207 functions as a critical transcription factor by transcriptionally regulating the expression of the pluripotency factor OCT4 (Fang et al, 2018), thereby being implicated in the maintenance of self-renewal and pluripotency. Likewise, ZNF207 has been shown to enhance reprogramming efficiency towards pluripotency (Toh et al, 2016). Alternative splicing (AS), in which splice sites in primary transcripts are differentially selected to produce structurally and functionally distinct mRNAs, plays a critical role in cell fate transitions, development, and disease (Gabut et al, 2011). ZNF207 undergoes AS during somatic cell reprogramming and differentiation of human ESCs, an isoform switch that seems to be required for the generation of induced pluripotent stem cells (iPSCs), and it might also be necessary to maintain ESC self-renewal and to induce proper differentiation programs (Toh et al, 2016; Fang et al, 2018).
Here, we show that ZFP207 plays an important role in the control of mouse ESC identity by a mechanism that differs from the one observed in human ESCs. Specifically, in mouse ESCs, ZFP207 does not regulate Oct4 transcription but increases OCT4 protein stability by disrupting ubiquitin-dependent proteasomal degradation. In addition, depletion of Zfp207 results in pluripotency defects and blocks neuroectodermal specification without significant changes in the transcriptome of stem cell and neural genes. ZFP207 regulates the expression of the spliceosome, and silencing of Zfp207 leads to aberrant AS patterns. We further describe ZFP207 as a novel RNA-binding protein (RBP), which might directly affect RNA fate. Taken together, this study uncovers the versatile species-specific roles of ZFP207 and the link to co- and post-transcriptional pathways that impact cell-fate decisions of mouse ESCs.
Results Silencing of Zfp207 impairs proliferation and mouse ESC identityTo explore the function of ZFP207 in mouse ESCs, we analyzed the expression of Zfp207 in retinoic acid (RA)-induced differentiation towards the neural lineage (Fig 1A) and in spontaneous differentiation of ESCs into the three germ layers by embryoid body (EB) formation (Fig 1B). Expression of the neuronal differentiation marker Nestin (Fig 1A) and of the pluripotency factor Oct4 (also known as Pou5f1; Fig 1A and B) was used to monitor proper cell differentiation. Real-time quantitative reverse transcription PCR (RT-qPCR) revealed that Zfp207 was significantly enriched in ESCs compared to differentiated cells (Fig 1A and B), and its expression levels gradually decreased along the course of differentiation, correlating with the decrease of ZFP207 and OCT4 protein levels (Fig 1C and D).

Figure 1. Depletion of Zfp207 leads to growth defects of mouse ESCs
A, B. RT-qPCR analysis of Zfp207 and Oct4 in mouse ESCs during (A) retinoic acid (RA)-induced and (B) embryoid body (EB)-mediated differentiation. Nestin was used as a neuronal differentiation marker to monitor RA-mediated differentiation. mRNA levels are relative to the expression at day 0. C, D. Western blot of ZFP207 and OCT4 of mouse ESCs during (C) RA-induced and (D) EB-mediated differentiation. E, F. (E) RT-qPCR and (F) western blot to monitor the knockdown efficiency of Zfp207 (sh1 and sh2). G. Cell proliferation rate of shScr, sh1, and sh2 ESCs assessed over a period of 6 days. H. Percentage of live (Annexin V−) and apoptotic cells (Annexin V+) in sh1 and sh2 mouse ESCs compared to shScr. I. Bar chart displaying the cell cycle distribution in sh1 and sh2 mouse ESCs relative to shScr. J, K. (J) AP staining of shScr and Zfp207-depleted (sh1 and sh2) mouse ESCs. Scale bars, 20 µM. (K) Percentage of fully differentiated (FD), partially differentiated (PD) and undifferentiated (UN) ESC colonies in shScr, sh1 and sh2. L. Immunofluorescence analysis of SSEA1 in shScr, sh1, and sh2 ESCs. DAPI was used as the nuclear marker. Scale bars, 20 μm. M, N. (M) Western blot of ZFP207, OCT4, NANOG, and SOX2 in shScr, sh1 and sh2 ESCs and (N) RT-qPCR analysis of Oct4, Nanog, and Sox2 in shScr, sh1 and sh2 ESCs; data is relative to shScr. O, P. (O) Western blot of OCT4 during a 9-h cycloheximide (CHX) time course treatment in shScr, sh1 and sh2 ESCs. (P) Protein degradation curves were made after quantification and normalization of the bands from (O). Q, R. (Q) Western blot of OCT4 during 4 and 8 h treatment with the proteasome inhibitor MG132. (R) graph bars after quantification and normalization of the bands from (Q).Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. §P < 0.05, §§P < 0.01, §§§P < 0.001 (D0 versus other indicated days). *P < 0.05, **P < 0.01, ***P < 0.001, n.s = no significant (shScr versus sh1 or sh2). A, B, G, H and I: unpaired Student’s t-test; E, N, P and R: ordinary one-way ANOVA.
Source data are available online for this figure.
To better understand the role of ZFP207 in pluripotency and differentiation, we aimed to generate CRISPR/Cas9-mediated knockout (KO) of Zfp207 in mouse ESCs. We used two different strategies: (i) one single-guide RNA (sgRNAs #1) targeting exon 3; and (ii) two distinct single-guide RNAs (#1 and #2) targeting the region containing exon 3 and exon 9, which included the microtubule-binding region domain of ZFP207 (Appendix Fig S1A). After picking and expanding individual clones, Western blot analysis and subsequent clone genotyping confirmed that all clones analyzed (94) displayed heterozygosity for Zfp207 (Appendix Fig S1B–E) suggesting that homozygous deletion of ZFP207 is lethal as previously suggested (Blomen et al, 2015). Next, we conducted loss-of-function assays by using two distinct short-hairpin RNAs (shRNAs) against Zfp207 (thereafter referred as knockdown 1 and 2 (KD1 and KD2)), to ensure that the observed phenotype is due to Zfp207 depletion and not due to shRNA off-target effects. Silencing of Zfp207 gene expression in both KD1 and KD2 ESCs was confirmed by RT-qPCR (Fig 1E) and by Western blot (Fig 1F). Depletion of Zfp207 led to a reduced proliferation capacity compared to ESCs transduced with scrambled shRNA (thereafter referred as control), although to a lesser degree in KD1 than in KD2 ESCs (Fig 1G; 1.3 and 2-fold, respectively). In addition, KD1 and KD2 ESCs showed a significant 1.5-fold increased apoptotic rate compared to control ESCs (Fig 1H) while no significant differences in the cell cycle profile were detected between the three cell lines (Fig 1I). Overall, our results indicate that ZFP207 is required for the proper proliferation of mouse ESCs.
Zfp207-depleted colonies displayed the typical morphology of differentiating ESCs with flat appearance and undefined colony borders (Fig 1J). Consistently, we detected reduced metabolic activity in ESC after depletion of Zfp207 determined by alkaline phosphatase (AP) activity assay (Fig 1J). Specifically, downregulation of Zfp207 resulted in a significant increase in the percentage of partially differentiated colonies, whereas the percentage of undifferentiated colonies was significantly decreased compared to control ESCs (Fig 1K). Such increase in the percentage of partially and finally differentiated colonies in KD1 and KD2 ESCs is a consequence of impaired ESC function as immunofluorescence analysis revealed that silencing of Zfp207 leads to a decrease of the pluripotency surface marker SSEA1 (Fig 1L).
Since ZFP207 regulates self-renewal and pluripotency in human ESCs, we next analyzed the expression of the core pluripotency factors and observed that Zfp207 KD1 and KD2 ESCs expressed lower OCT4, but not NANOG and SOX2, compared to control ESCs (Fig 1M). Interestingly, mRNA levels of the three pluripotency factors, including those of Oct4 (Fig 1N), were unaltered upon silencing of Zfp207, indicating that ZFP207 could regulate the expression of OCT4 post-transcriptionally. To test this hypothesis, we treated KD1 and KD2 as well as control ESCs with the protein synthesis inhibitor cycloheximide (CHX). Silencing of Zfp207 led to a decrease in the half-life of OCT4 (Fig 1O and P), suggesting that ZFP207 promotes the stability of this pluripotency factor. In addition, treatment with the proteasome inhibitor MG132 restored OCT4 protein levels in mouse ESCs depleted of Zfp207 (Fig 1Q and R), indicating that ZFP207 interferes with the turnover of this pluripotency factor.
To assess whether ZFP207 sustains self-renewal in the naïve ground state, we cultured mouse ESCs in the presence of kinase inhibitors against MAP kinase (MEK) and glycogen synthase kinase 3β (GSK-3β; “2i”) and leukemia inhibitory factor (LIF) (2iL medium) (Martello & Smith, 2014). Although the number of partially differentiated colonies in 2iL-cultured ESCs increased upon silencing of Zfp207 assessed by a loss of reactivity to AP (Fig EV1A and B), RNA and protein levels of OCT4, NANOG, and SOX2 were unaffected (Fig EV1C and D). Likewise, no changes in SSEA1 staining were observed in 2iL-cultured ESCs upon Zfp207 knockdown (Fig EV1E), suggesting that ZFP207 is dispensable for the maintenance of pluripotency in the ground state. Therefore, we performed the experiments in mouse ESCs cultured in conventional medium containing serum and LIF, that is, metastable ESCs, where we observed that ZFP207 contributes to stem cell identity by regulating post-transcriptional networks.
Click here to expand this figure.
Figure EV1. ZFP207 is dispensable in mouse ESCs maintained in ground state pluripotency
A, B. (A) AP staining of shScr and Zfp207-depleted (sh1 and sh2) mouse ESCs cultured in 2iL. Scale bars, 20 µM. (B) Percentage of fully differentiated (FD), partially differentiated (PD) and undifferentiated (UN) ESC colonies in shScr, sh1 and sh2 cultured in 2iL. C, D. (C) Western blot of ZFP207, OCT4, NANOG, and SOX2 and (D) RT-qPCR analysis of Zfp207,Oct4, Nanog, and Sox2 in shScr, sh1, and sh2 ESCs cultured in 2iL; data are relative to shScr. E. Immunofluorescence analysis of SSEA1 in shScr, sh1, and sh2 ESCs cultured in 2iL. DAPI was used as the nuclear marker. Scale bars, 20 μm.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. **P < 0.01, n.s = no significant difference (shScr versus sh1 or sh2). B: unpaired Student’s t-test; D: Ordinary one-way ANOVA.
Source data are available online for this figure.
ZFP207 is required for proper EB differentiationWe next interrogated the role of ZFP207 during lineage specification by assessing the potential of KD1, KD2, and control mouse ESCs to spontaneously differentiate into EBs recapitulating early mouse embryo development. Zfp207 KD1 and KD2 ESCs were able to form EBs (Fig 2A). Although the cells remained as solid aggregates, we observed a decrease in the size of both Zfp207 KD1 and KD2 EBs compared to control EBs (Fig 2B), suggesting intrinsic differences during the differentiation process among the different cell lines. Given that ZFP207 influenced the apoptotic rate of ESCs (Fig 1H), we sought to investigate the mechanisms underlying the reduced size of Zfp207 KD1 and KD2 compared to control EBs. Bromodeoxyuridine (BrdU) incorporation revealed that Zfp207 knockdown cells were associated with a significantly reduced growth rate compared with control EBs after day 4 of differentiation (Fig 2C and Appendix Fig S2A). However, we did not observe differences in apoptosis assessed by Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining performed along the course of differentiation or Annexin V staining of EBs at day 8 (Appendix Fig S2B–D). Altogether these results suggest that, albeit ZFP207 is critical for proper EB proliferation, its expression is not necessary for the viability of the EBs.

Figure 2. Loss of Zfp207 results in defective differentiation
A, B. (A) Bright-field images (10× (left) and 4× (right) magnification) and (B) quantification of embryoid bodies (EB) generated from shScr, sh1, and sh2 at day 8 of differentiation. Scale bars, 200 µM. C. Quantification of BrdU incorporation in shScr, sh1, and sh2 ESCs at the indicated days of EB differentiation. Data are relative to shScr. D. RT-qPCR of Zfp207, the pluripotency genes (Oct4, Nanog and Sox2), the mesodermal markers (Msx1 and Brachyury (T)), the endodermal markers (Foxa2, Sox17), and the neural-associated genes (Pax6, Sox11, and Nestin) in shScr, sh1, and sh2 ESCs along the time-course of EB-mediated differentiation. mRNA levels are relative to the expression of shScr at day 0.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (shScr versus sh1 or sh2). B, C: unpaired Student’s t-test; D: ratio paired Student’s t-test.
As expected, while Zfp207 expression was gradually downregulated during the course of differentiation in control cells, Zfp207 mRNA levels remained low in both KD1 and KD2 cells (Fig 2D). The expression of the pluripotent genes Nanog and Sox2 decreased rapidly in all the cell lines, whereas Oct4 decreased progressively, being its expression higher at the latest stage of the time course differentiation in EBs depleted of Zfp207 compared to control (Fig 2D). EBs originated from the three shRNA-infected ESCs showed normal levels of the mesodermal (Msx1 and Brachyury or T) and endodermal (Foxa2 and Sox17) lineage-specific markers (Fig 2D). However, silencing of Zfp207 impaired ectodermal specification as shown by decreased expression of Pax6 and Sox11 at day 4, and Nestin during the course of differentiation in Zfp207 KD EBs compared to controls (Fig 2D and Appendix Fig S2E).
Silencing of Zfp207 blocks differentiation of ESCs to neural progenitor cells and subsequently to neuronsTo investigate whether ZFP207 plays a role in neurogenesis, KD1, KD2, and control ESCs were subjected to neural differentiation (Fig 3A). Albeit the distinct cell lines did not display major morphological differences at early stages of differentiation (when growing as EBs), KD1 and KD2 underwent a complete block of neuronal differentiation potential at day five (Appendix Fig S3A), and we were not able to characterize Zfp207 KD1 and KD2 cells at later stages as they died at day 6. However, in control cell lines, NESTIN- and TUJ1-immunopositive neuronal projections appeared at day 8 and were kept in culture until day twelve (Fig 3B). Hence, we sought to also assess the viability and proliferation phenotype of Zfp207-depleted mouse ESCs upon neural differentiation. Immunostaining of TUNEL and cleaved caspase 3 revealed that a large proportion of Zfp207 knockdown cells underwent apoptosis during differentiation (Fig 3C–E, Appendix Fig S3B and C). In addition, Zfp207 depletion dramatically decreased cell proliferation at day 4 of differentiation as measured by BrdU incorporation (Fig 3F), indicating that ZFP207 play important roles in maintaining neural stem/progenitor cell properties, including their survival and proliferative capacities.

Figure 3. ZFP207 is essential for neural cell fate specification
A. Schematic depiction for the neuroectodermal-directed differentiation. DMEM/F12 supplemented with N2B27 and retinoic acid (RA) was added after two days of EB culture. B. Immunostaining of NESTIN (green) and TUJ1 (red) of neural progenitors generated from shScr on day 8 and 10 of the neuroectodermal differentiation. Nuclei were counterstained with DAPI. Scale bar, 20 μM. C–E. (C) TUNEL (green), (D) CASPASE 3 (red) staining and (E) quantification of the signal in shScr, sh1, and sh2 at day 5 of neuroectodermal differentiation. Nuclei were counterstained with DAPI. Scale bar, 20 µM. F. Flow cytometric profile and quantification of BrdU incorporation at day 4 of neuroectodermal differentiation. G. RT-qPCR of Zfp207 and neural-associated markers in shScr, sh1, and sh2 along the course of neural differentiation. mRNA levels are relative to shScr at day 0.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (shScr versus sh1 or sh2). E, F, and G: unpaired Student’s t-test.
Similar to RA- and EB-mediated differentiation, Zfp207 expression was downregulated during the course of neurogenesis in control cells, and Zfp207 mRNA levels remained low in both KD1 and KD2 cells (Fig 3G). We also monitored progression of ESC differentiation toward neural fates by RT-qPCR analysis of neuroectodermal markers. Zfp207-depleted cell lines failed to activate the expression of Pax6, Sox11, and Nestin (Fig 3G and Appendix Fig S3D), suggesting a stall at early ectodermal differentiation. No defects were found in the expression of Tuj1 at the transcriptional level as the upregulation of this neuronal marker occurred at later stages when the KD1 and KD2 cells had died (Fig 3G).
We next assessed whether ZFP207 is required for early or terminal differentiation of neurons by derivation of neural progenitor cells (NPCs) from KD1, KD2, and control ESCs (Hanafiah et al, 2020) (Fig EV2A). Even though Zfp207 knockdown mouse ESCs were successfully differentiated to EBs (day 4), it was immediately apparent that KD1 and KD2 NPCs were smaller and all died after day 6 (Fig EV2B and C). In addition, RT-qPCR analysis indicated that KD1 and KD2 failed to activate the expression of ectodermal markers, again consistent with a blockade at early ectodermal differentiation upon ZFP207 deficiency (Fig EV2D).
Click here to expand this figure.
Figure EV2. Zfp207 knockdown ESCs cannot generate NPCs
A. Schematic diagram for neural progenitor cells (NPC) generation. Retinoic acid (RA) was added after four days of EB culture. B, C. (B) Representative bright-field images (10×) and (C) quantification of NPCs on day 6 of differentiation. Scale bars, 200 μm. D. RT-qPCR of Zfp207 and neural-associated markers in shScr, sh1 and sh2 along the course of neural progenitor (NPC) differentiation. mRNA levels are relative to shScr at day 0.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. *P < 0.05, **P < 0.01, ****P < 0.0001. C: Ordinary one-way ANOVA; D: two-way ANOVA.
To demonstrate the specificity of the phenotype observed upon silencing of Zfp207, we engineered a tet(ON)-ZFP207 cell line in which endogenous Zfp207 can be depleted by shRNA2 and replaced with a shRNA2-immune exogenous Zfp207 cDNA responsive to doxycycline (Dox) treatment. As expected, Dox treatment in tet(ON)-ZFP207 KD2 induced the expression of ZFP207 which resulted in an increase of OCT4 protein levels but not Oct4 mRNA (Fig EV3A and B). In addition, re-expression of ZFP207 rescued the proliferation and apoptotic defect of tet(ON)-ZFP207 KD2 (Fig EV3C and D). Furthermore, upon addition of Dox, tet(ON)-ZFP207 KD2 displayed a characteristic ESC-like morphology, and the number of partially differentiated colonies was increased whereas the percentage of final differentiated colonies was significantly decreased compared to Zfp207 KD2 ESCs without Dox induction (Fig EV3E and F). Strikingly, re-expression of ZFP207 was not sufficient to return the normal number of undifferentiated colonies (Fig EV3E and F). Nevertheless, the levels of SSEA1 staining of tet(ON)-ZFP207 KD2 upon Dox treatment were comparable to control ESCs (Fig EV3G), confirming that ZFP207 is critical for the maintenance of mouse ESCs.
Click here to expand this figure.
Figure EV3. Re-expression of Zfp207 in tet(ON)-ZFP207 KD2 rescues the ESC phenotype but it is not sufficient to fully differentiate tet(ON)-ZFP207 KD2 to neurons
A, B. (A) Western blot of ZFP207 and OCT4 and (B) RT-qPCR of Zfp207 and Oct4 in the tet(ON)-ZFP207 cell line subjected to shScr or sh2 in the absence (−) or presence (+) of doxycycline (Dox) as indicated. mRNA levels are relative to the expression in shScr. C. Relative proliferation rate of tet(ON)-ZFP207 ESCs with shScr and sh2 −/+ Dox assessed over a period of 8 days. D. Percentage of live (Annexin V−) and apoptotic cells (Annexin V+) in tet(ON)-ZFP207 ESCs with shScr and sh2 −/+ Dox. E, F. (E) AP staining of shScr and sh2 −/+ Dox in tet(ON)-ZFP207 ESCs. Scale bar, 50 µM. (F) Percentage of fully differentiated (FD), partially differentiated (PD) and undifferentiated (UN) ESC colonies in shScr and sh2 −/+ Dox treatment in tet(ON)-ZFP207. G. Immunofluorescence analysis of SSEA1 in tet(ON)-ZFP207 ESCs with shScr and sh2 −/+ Dox. DAPI was used as the nuclear marker. Scale bars, 20 μm. H. Representative bright-field images (20x) and quantification of neurospheres on day 4 of differentiation. Scale bars, 200 μm. I. RT-qPCR of neural-associated markers in tet(ON)-ZFP207 ESCs with shScr and sh2 −/+ Dox at day 4 (upper panel) and tet(ON)-ZFP207 ESCs with shScr and sh2 +Dox at day 5 (lower panel). mRNA levels are relative to shScr. J. TUNEL (green) staining in tet(ON)-ZFP207 ESCs with shScr and sh2 +Dox at day 5. Nuclei were counterstained with DAPI. Scale bar, 20 µm.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns = no significant difference (sh2 versus sh2 Dox). B, C, D, F and I: unpaired Student’s t-test.
Source data are available online for this figure.
We next investigated whether the neuroectoderm blockade was specific to the depletion of Zfp207 by subjecting the tet(ON)-ZFP207 cell line to neural differentiation. Of note, tet(ON)-ZFP207 KD2 cells died at day 4; therefore, it was not possible to characterize the phenotype at later days of differentiation. However, after Dox treatment, tet(ON)-ZFP207 KD2 regained the ability to differentiate being the size of the neurospheres similar to that of the control cells (EV3H). In addition, re-expression of ZFP207 restored the ability to induce the expression of Pax6, Sox11, and Nestin, especially at day 5, where no differences in the expression of the neuroectodermal markers were detected between control and tet(ON)-ZFP207 KD2 induced with Dox (EV3I). However, the blockade phenotype was only partially recovered as rescue cells died at later stages and displayed increase TUNEL staining (EV3J), illustrating a complex regulatory interplay of Zfp207 isoforms occurring during neural differentiation that will be discussed later.
ZFP207 does not transcriptionally regulate the ectodermal lineageIn order to gain insight on the role of ZFP207 in ESC pluripotency, we analyzed the global transcriptome response to Zfp207 depletion (Fig 4A). RNA-sequencing (RNA-seq) analysis identified 382 and 522 genes that were downregulated in KD1 and KD2 ESCs, respectively (fold change > 1.5; P < 0.05; Fig 4B and Dataset EV1). The differences in the number of downregulated genes between the two KDs could result in the more severe phenotype observed in cells transduced with shRNA2 compared to shRNA1 (Fig 1G). The effect on upregulation was more robust, whereby 1,062 and 1,082 genes were upregulated in KD1 and KD2 ESCs, respectively, compared to control ESCs (Fig 4B). Gene ontology (GO) analysis of biological processes of common downregulated genes revealed generic functions, which included mitotic sister chromatid segregation among other categories (Fig 4C). Strikingly, poly(A)+ RNA binding was among the most represented molecular function GO categories (Appendix Fig S4A). According to the reported function of ZFP207 in kinetochore–microtubule attachment (Dai et al, 2016), top GO categories for molecular function of common downregulated genes also included microtubule binding (Appendix Fig S4A). We validated these results by performing RT-qPCR analysis of downregulated mitotic sister chromatid segregation genes (e.g., Cdca8 and Cep57l1; Appendix Fig S4B). Upregulated genes were associated with GO biological processes related to RNA splicing and processing, and positive regulation of transcription (Fig 4D). Similar to the common downregulated genes, RNA binding, including poly(A)+ and mRNA binding, were among the most represented categories of GO molecular function (Appendix Fig S4C), suggesting a role of ZFP207 in controlling the expression of putative RBPs, primarily involved in RNA splicing and mRNA processing (Appendix Fig S4D).

Figure 4. The neural transcriptome is not altered upon depletion of Zfp207
A, B. (A) Volcano plots of common differentially expressed genes in shScr and Zfp207-depleted (sh1 and sh2) mouse ESCs. Upregulated (Up) and downregulated (Down) genes are indicated in red and blue, respectively (P < 0.05; > 1.5-fold). Zfp207 is depicted in black. Grey dots indicate non-significant (NS) and < 1.5-fold differential expressed genes. (B) Venn diagram depicting the overlap of downregulated and upregulated genes between sh1 and sh2 ESCs (FC > 1.5 and P < 0.05). FDR value was calculated with the Benjamini–Hochberg correction. C, D. Gene ontology (GO) analysis of biological processes associated with common (C) downregulated and (D) upregulated genes in Zfp207-depleted ESCs (sh1 and sh2); NHEJ: Nonhomologous end joining. E–H. Heatmap of column z-scores of log2 transformed values of genes related to (E) pluripotency and (G) ectoderm in shScr, sh1, and sh2 ESCs. GSEA plots depicting the expression of (F) ESC-related and (H) ectodermal genes in Zfp207-depleted ESCs compared to shScr. High and low expression of genes is represented in red and blue color, respectively. FDR, false discovery rate; NES, normalized enrichment score.Depletion of Zfp207 did not lead to aberrant transcriptional programs that could explain the developmental defects. Hence, there were no differences in the expression of ESC genes upon silencing of Zfp207 (Fig 4E and F). In addition, we did not find major transcriptional differences in genes associated with the neural lineage that could reflect the stall at early ectodermal differentiation in KD1 and KD2 ESCs (Fig 4G and H). This is in striking contrast to what has been reported in human ESCs where depletion of ZNF207 impairs neuroectodermal specification by transcriptionally regulating the expression of genes associated with the ectoderm lineage (Fang et al, 2018). Noteworthy, in human ESCs, the isoforms A and C of ZNF207, retaining the exon 9, are highly abundant compared to mouse ESCs, where the isoforms 1 and 2 of Zfp207, retaining the aforementioned exon 9, are highly expressed in differentiated cells (Fig EV4A and B). Hence, in both species, there is an antagonistic switch toward using different isoforms during differentiation, which could partially explain the differences observed between mouse and human ESCs. Noteworthy, Zfp207 isoform switching occurring during differentiation could also explain why re-expression of one isoform of Zfp207 in the rescue experiments, that is, isoform 3, which is highly expressed in mouse ESCs, is not sufficient to fully rescue the phenotype at later stages of differentiation (Fig EV3H and I). As ZNF207 is a downstream splicing target of SFRS11 during somatic cell reprogramming (Toh et al, 2016), we sought to analyze whether this splicing factor was also modulating mouse Zfp207 splicing. Similar to what is observed in the human ortholog, depletion of Sfrs11 increased retention of exon 9, corresponding to the isoforms 1 and 2 of Zfp207 (Fig EV4, EV5). Therefore, SFRS11 modulates the same splicing event in mouse and human ESCs but with divergent consequences.
Click here to expand this figure.
Figure EV4. Zfp207 undergoes alternative splicing during differentiationz
A, B. (A) Schema showing a segment in transcript variants with alternatively spliced exons of Zfp207 in mouse and human (left panel). The differential switch of the splice forms between the ESC state and neural-directed differentiation in mouse and human are also depicted (right panel). (B) RT-PCR analysis of the AS forms of Zfp207 at the indicated time points of retinoic acid (RA) -induced differentiation, embryoid body (EB) generation and neuroectodermal (NE) differentiation of mouse ESCs. C–E. (C) RT-qPCR and (D) western blot to monitor the knockdown efficiency of Sfrs11 (sh1 and sh2). (E) RT-PCR analysis of the AS forms of Zfp207 upon depletion of Sfrs11.Data information: Data are presented as mean ± SEM or representative images of n ≥ 3 independent biological experiments. ***P < 0.001, ****P < 0.0001 (shScr versus sh1 or sh2). C: unpaired Student’s t-test.
Source data are available online for this figure.
Click here to expand this figure.
Figure EV5. Alternative splicing switches in Zfp207 knockdown ESCs are prevalent in differentiated cells
Schema depicting the types of AS events: SE, skipped exon; MX, mutually exclusive exons; A5, alternative 5′ splice-site; A3, alternative 3′ splice site; RI, retained intron; AF, alternative first exon; AL, alternative last exon, generated by the software SUPPA2 with the specific coordinates, including start (s) and end (e). The form of the AS event is depicted in black. Pie chart showing the AS events types distribution. Volcano plot depicting the correlation between gene expression levels and dPSI index resulted from RNA-seq data analysis in control (scShr) and Zfp207 depleted mouse ESCs (sh1 + sh2). Gene expression levels are presented as log2 transformed values. Differential percent spliced in index (dPSI) range is between −1 and +1. Gene ontology enrichment analysis with Bioplanet software for genes undergoing different splicing events upon Zfp207 depletion (sh1 and sh2). Representative RT-PCR analysis of AS events upon neural differentiation in mouse ESCs for Mta1 (AS1 and AS2), Tbx3, Hnrnp1a, Nasp, Myef2, Fxr1, and Eif4H. The structure of each isoform is indicated (not to scale). Alternative exons are blue. PSI was quantified for each condition. ZFP207 regulates alternative splicing in mouse ESCsAS is a fundamental process that increases proteomic diversity in a species-manner but not organ-specific manner (Barbosa-Morais et al, 2012), and it also plays a central role in the regulation of ESC-specific transcriptional programs (Gabut et al, 2011). ZNF207 has been shown to influence pre-mRNA splicing in cancer cells (Wan et al, 2015). Consistent with this observation, our GO enrichment analysis of genes upregulated upon Zfp207 silencing revealed significant enrichment of GO terms related to RNA splicing (Figs 4D and 5A), and some randomly selected spliceosomal genes (Mbnl2, Rbm3, and Sf3b3) were further validated by RT-qPCR (Fig 5B).

Figure 5. Silencing of Zfp207 leads to AS defects
A. Heatmap of column z-scores of log2 transformed values of genes involved in alternative splicing (AS) in shScr, sh1 and sh2 mouse ESCs. B. RT-qPCR analysis of Rbm3, Mbnl2, and Sf3b3 in shScr, sh1 and sh2 ESCs. mRNA expression is relative to shScr and presented as mean ± SEM of n ≥ 3 independent biological experiments; *P < 0.05, **P < 0.01, ***P < 0.001 (shScr versus sh1 or sh2); unpaired Student’s t-test. C. Violin plot depicting statistically significant spliced in (PSI) events in each category in sh1 and sh2 related to shScr. Alternative 3′ (A3); Alternative 5′ (A5); Alternative first exon (AF); Alternative last exon (AL); Mutually exclusive exons (MX); Intron retention (RI); Exon skipping (SE). The dotted line indicates ΔPSI = 0. D, E. (D) Venn diagram showing the overlap of AS in sh1 and sh2. (E) Gene ontology analysis of common genes undergoing AS in sh1 and sh2 mouse ESCs according to the type of AS event. F. RT-PCR of Zfp207-regulated AS events. For Mta1, AS1 (alternative splicing 1) and AS2 (alternative splicing 2). #1 and #2 indicate distinct biological replicates. *Denotes an isoform that was not taken in consideration for the quantification. The structure of each isoform is indicated (not to scale). Alternative exons are blue. The percent spliced in (PSI) was quantified for each condition.We therefore sought to investigate whether ZFP207 could control ESC function by a splicing-related mechanism. To this end, reads from RNA-seq data were mapped to exon-splice junction’s sites in order to elucidate genome-wide differential AS events (DSEs) (Alamancos et al, 2015; Trincado et al,
留言 (0)