
記住我




Multiple myeloma (MM) is a malignant disorder characterized by the uncontrolled expansion of clonal plasma cells in the bone marrow, eventually leading to organ dysfunction and death (Laubach et al, 2011). The global burden of MM has increased by 126% over the past 30 years, with highest incidences in developed countries (Cowan et al, 2018). The introduction of proteasome inhibitors (PIs) like bortezomib (BZ) in MM chemotherapies has revolutionized MM management, and combinations of BZ with immunomodulators (IMiDs) and dexamethasone have become the gold standard for MM treatment (Kouroukis et al, 2014). However, BZ can induce severe side effects such as peripheral neuropathy, requiring discontinuation of therapy (Lawasut et al, 2012). Moreover, many patients develop resistance to BZ whose molecular basis remains poorly understood, and less than half of them survive beyond 5 years post-diagnosis. Second generation PIs were recently developed to treat patients who are resistant or intolerant to BZ (Gandolfi et al, 2017), but these drugs induce other types of side effects and do not prevent relapse. Therefore, despite significant therapeutic advances, MM remains an incurable disease and the identification of new therapeutic targets is critically needed.
The clinical efficacy of BZ is primarily attributed to its ability to induce the accumulation of misfolded proteins including immunoglobulins in the cytoplasm of MM cells, leading to lethal proteotoxic stress (Gandolfi et al, 2017). BZ also alters the survival and proliferation of MM cells in several other ways, such as inhibition of NF-κB oncogenic signaling, suppression of pro-adhesive cross-talks with bone marrow stromal cells, and prevention of growth stimulation by cytokines like interleukin (IL)-6 (Mahindra et al, 2010; Rosean et al, 2014). Among the different strategies that are currently explored to complement or replace PIs in MM chemotherapies are novel means to disrupt the homeostatic regulation of the secretory apparatus via proteolytic routes or endoplasmic reticulum (ER)-associated protein degradation (ERAD) components other than the proteasome (Auner & Cenci, 2015). Despite its central importance in MM cell biology, the interest of targeting immunoglobulin transport into the ER has not yet been explored due to the lack of suitable inhibitor.
We reported recently that mycolactone, a diffusible lipid toxin secreted by the human pathogen Mycobacterium ulcerans, operates by inhibiting the mammalian translocon (Sec61) (Baron et al, 2016; Demangel & High, 2018). By targeting the central pore-forming subunit of Sec61 (Sec61α), mycolactone prevents the import of newly synthesized secreted and transmembrane proteins into the ER, leading to their cytosolic degradation by the proteasome (Hall et al, 2014; McKenna et al, 2016, 2017). Contrary to the Sec61 inhibitor cotransin (Mackinnon et al, 2014), mycolactone is not substrate selective and blocks the translocation of the vast majority of Sec61 clients (Baron et al, 2016; Morel et al, 2018). The only substrates resisting its inhibitory action are transmembrane proteins belonging to the rare Type III subgroup (McKenna et al, 2016, 2017; Demangel & High, 2018; Morel et al, 2018). Within 1 h of treatment, mycolactone-treated cells become defective for production of most secreted proteins and membrane-anchored receptors. If sustained, mycolactone treatment triggers proteotoxic stress responses in cytosol and ER, ultimately inducing apoptosis (Morel et al, 2018; Ogbechi et al, 2018). Notably, a point mutation (R66G) in the Sec61α amino acid sequences preventing mycolactone binding without affecting the translocon functionality was sufficient to prevent stress responses and associated cytotoxicity, demonstrating that Sec61 inhibition by mycolactone is the molecular mechanism driving cell death (Baron et al, 2016).
Based on these findings, we hypothesized that Sec61 blockade may suppress survival and growth of MM cells in two ways: by preventing the expression of membrane receptors that are key to MM cell division and dissemination, and by generating lethal proteotoxic stress. The present study uses mycolactone as a prototypical Sec61 blocker to establish proof of concept, and evaluates the translational potential of Sec61 inhibitors in the treatment of MM.
Results Sec61 blockade by mycolactone alters the biology and viability of MM cell linesTo assess the effect of mycolactone on MM cell viability, three human cell lines (MM.1S, JIM3, and KMS-11) were treated with increasing concentrations of mycolactone for 24–72 h, and the induction of cell apoptosis was monitored by phosphatidylserine exposure (Annexin V staining) and loss of membrane integrity (PI staining) (Appendix Fig S1). While the three cell lines differed in sensitivity to mycolactone, MM.1S being the most resistant, a dose- and time-dependent induction of apoptotic cell death was consistently observed after 48 h of treatment (Fig 1A). In all cell lines, initiation of apoptosis was preceded by a decrease in surface expression of the MM cell marker CD38 (Van De Donk et al, 2018) (Fig 1B). The plasma cell marker CD138 supports MM cell survival in the bone marrow by promoting growth factor signaling (Akhmetzyanova et al, 2020). While not detected on JIM3 and KMS-11, CD138 expression by MM.1S was also dose dependently decreased by mycolactone after 24 h (Fig 1B). IL-6 receptor (IL-6R) and CD40 are two other MM cell markers whose signaling play a crucial role in MM development and dissemination (Tai et al, 2005; Rosean et al, 2014). When expressed by the MM cell lines, these receptors were also rapidly and potently downregulated by mycolactone treatment (Fig 1B).

Figure 1. Sec61 blockade by mycolactone strongly alters the biology and viability of MM cell lines
A. Effect of mycolactone on MM.1S, JIM3, and KMS-11 cell viability after 48 and 72 h of treatment, as measured by exposure of Annexin V and incorporation of PI. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, gated as in Appendix Fig S1, relative to total cells. B, C. Inhibitory effect of mycolactone on cell expression of CD38 (Type II) and CD138, IL-6R, and CD40 (Type I) transmembrane proteins (B). Stimulatory effect of mycolactone on cell expression of BCMA, a Type III transmembrane protein (C). Mean fluorescence intensities (MFIs) were measured in live cells 48 h after addition of mycolactone. Data are Mean MFIs ± SD from technical duplicates of two independent experiments, relative to vehicle-treated controls (Ctrl). D. Inhibitory effect of mycolactone on immunoglobulin (lambda chain) secretion, as quantified by ELISA in culture supernatants after 24 h of treatment. Data are Mean concentrations ± SD of secreted lambda chain (µg/ml) from technical duplicates of two independent experiments (MM.1S) or Mean of secreted lambda chain concentrations (µg/ml) from technical duplicates (JIM3 and KMS-11), relative to vehicle-treated controls. E. Correlation between immunoglobulin lambda chain (Ig) secretion after 24 h (D) and MM cell line mortality after 72 h (A), in the three cell lines treated with 50 nM mycolactone. Slopes (R) and statistical significance (P values) are indicated.Data information: In A (all cell lines) and D (JIM3 and KMS-11), shown data are representative of two independent experiments with similar results.
In contrast to CD38, CD138, IL-6R, and CD40, all Type I or II transmembrane proteins efficiently blocked in translocation by mycolactone; Type III transmembrane proteins are Sec61 substrates resisting mycolactone inhibition (Baron et al, 2016; McKenna et al, 2016, 2017; Morel et al, 2018). B-cell maturation antigen (BCMA) is a Type III protein that is typically expressed by MM cells and the target of next-generation immunotherapies (Cho et al, 2020). In all cell lines, mycolactone dose dependently increased cell surface expression of BCMA (Fig 1C). On the opposite, secreted proteins are Sec61 substrates that are efficiently blocked in translocation by mycolactone (Baron et al, 2016; McKenna et al, 2016, 2017; Morel et al, 2018). Figure 1D shows that Sec61 inhibition efficiently decreased MM cell line secretion of immunoglobulin light chains after only 24 h of treatment, and this reduction closely correlated with the onset of cell death after 72 h (Fig 1E). In conclusion, Sec61 blockade by mycolactone induces programmed cell death in MM preceded by phenotypic defects in expression of Type I/II membrane receptors and secretion of immunoglobulins.
Mycolactone synergizes with proteasome inhibitors for induction of MM cell apoptosisThe clinical efficacy of BZ is believed to result from its capacity to induce unresolvable proteotoxic stress in MM cells via accumulation of misfolded proteins in the cytoplasm (Gandolfi et al, 2017). Since Sec61 substrates blocked in translocation by mycolactone are diverted to the proteasome for degradation (Hall et al, 2014), we reasoned that mycolactone may potentiate the anti-MM activity of BZ through generation of additional proteotoxic stress. While mycolactone-mediated cytotoxicity did not manifest before 48 h of treatment (Fig 1A), BZ induced significant mortality in all MM cell lines after only 24 h (Fig 2A). Cell lines nevertheless differed in susceptibility to BZ cytotoxicity, JIM3 and KMS-11 being relatively more resistant than MM.1S. To assess a potential synergy between mycolactone and BZ, MM cells were treated for 24 h with subtoxic doses of BZ together with increasing concentrations of mycolactone, and induction of apoptosis was assessed after 24 h. In all cell lines, the mycolactone–BZ combination induced more cell death than single drugs (Figs 2B and EV1). Heatmaps of synergy scores (Di Veroli et al, 2016) show that mycolactone synergized with BZ in all cell lines, irrespective of their basal resistance to BZ (Fig 2C). Similar findings were obtained with MM.1S cells treated with carfilzomib, a second-generation PI that is proposed to BZ-resistant MM patients (Fig 2D and E).

Figure 2. Mycolactone synergizes with PIs for induction of MM cell apoptosis
A. MM.1S, JIM3, and KMS-11 cells were incubated with increasing concentrations of BZ for 24 h. The proportion of live (Annexin V- PI-) cells was then measured by flow cytometry as in Appendix Fig S1. Data are Mean % of live cells from technical duplicates, relative to vehicle controls (Ctrl). B. MM.1S were treated for 24 h with mycolactone and/or BZ at the indicated concentrations. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, gated as in Appendix Fig S1, relative to total cells. C. Synergy between mycolactone and BZ in the MM.1S, JIM3, and KMS-11 cell lines, when treated as in (B). Data are Mean Loewe scores ± SD shown as heatmaps. N = 6 (cumulative data of three independent experiments with technical duplicates) for MM.1S and KMS-11, and N = 4 (cumulative data of two independent experiments with technical duplicates) for JIM3. Statistical significance was assessed by Student’s t-test: *P < 0.05; **P < 0.01; ***P < 0.001. D, E. MM.1S cell were treated as in (B) but using carfilzomib instead of BZ (D). Synergy scores (E) were calculated as in (C). Data are Mean Loewe scores of technical duplicates, shown as heatmaps.Data information: In A and B, shown data are representative of two and three independent experiments with similar results, respectively.
Click here to expand this figure.
Figure EV1. Mycolactone potentiates the effect of BZ in JIM3 and KMS-11
JIM3 and KMS-11 cells were treated with mycolactone and/or BZ at the indicated concentrations for 24 h. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, gated as in Appendix Fig S1, relative to total cells. They are representative of two independent experiments with similar results.
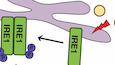
The mycolactone–BZ combination activates pro-apoptotic UPR in MM cellsWe next sought to determine if the synergistic induction of apoptosis in MM cell lines treated with mycolactone and BZ correlated with enhanced ER stress. The unfolded protein response (UPR) is activated by three ER-resident transmembrane sensor proteins: protein kinase RNA-like ER kinase (PERK), inositol requiring enzyme 1 (IRE1), and activating transcription factor (ATF) 6 (Fig 3A) (Hetz & Papa, 2018). Activation of PERK leads to the phosphorylation of eIF2α, with p-eIF2α reprogramming protein translation to promote the expression of stress response mRNAs such as ATF4. ATF4 subsequently drives the expression of genes alleviating stress such as the growth arrest and DNA damage-inducible protein (GADD34), which promotes p-eIF2α dephosphorylation in a negative feedback loop. Activation of IRE1 triggers the splicing of X-box-binding protein 1 (XBP1) mRNA into its transcriptionally active form (sXBP1), able to transcriptionally reprogram UPR target genes. Chronic stress promotes transition from adaptive to terminal UPR, marked by ATF4-mediated induction of pro-apoptotic C/EBP homology protein (CHOP) expression and accentuated by pro-death signals originating from IRE1 signaling. Activation of ATF6 leads to its cleavage into an active form in the Golgi, which contributes to upregulate CHOP.

Figure 3. The mycolactone–BZ combination activates pro-apoptotic UPR in MM.1S cells
A. Diagram illustrating the pro-apoptotic pathways activated by UPR sensors PERK, ATF6, and IRE1 upon accumulation of unfolded proteins in the ER lumen. B–G. In MM.1S cells treated with mycolactone and/or BZ at the indicated concentrations, or vehicle as control for 6 h: (B) ATF4 mRNA levels were quantified by qPCR; (C) ATF4 protein levels in cell lysates were assessed by Western blot (top panel) and quantified relatively to GADPH levels (lower panel); (D) CHOP and GADD34 mRNA levels were quantified by qPCR; (E) ATF6 and GAPDH protein levels in cell lysates were assessed by Western blot with molecular weight markers (kDa) indicated on the right; Bands corresponding to glycosylated (G-ATF6), non-glycosylated (NG-ATF6), and cleaved ATF6 are indicated by arrows; (F) Total RNA was isolated and used as a template for RT-PCR of XBP-1 (upper panel), which was then digested with Pst1 and separated on a 2% agarose gel (lower panel) with molecular weight markers (bp) indicated on the left; spliced XBP1 (sXBP1) mRNA levels were also quantified by qPCR and normalized to total (spliced + unspliced) XBP1 mRNA levels, and (G) BiP mRNA level was quantified by qPCR.Data information: Shown mRNA data are mean fold changes (2−ΔΔCT) ± SD, relative to untreated controls (cumulative data of at least two independent experiments with technical duplicates or triplicates), pairwise compared by nested one-way ANOVA with Tukey’s multiple-comparison test, exact P-values indicated. Thapsigargin (Tg, 2 µM, 6 h), tunicamycin (Tu, 2 µM, 6 h), or DTT (4 mM, 2 h) were used as positive controls. In C, E, and F, shown data are representative of two independent experiments with similar results.
In all MM cell lines treated with subcytotoxic concentrations of mycolactone and BZ for 6 h, we detected an increased expression of ATF4, at both the mRNA and protein levels, compared to single drug treatments (Figs 3B and C, and EV2A). Elevated ATF4 correlated with enhanced expression of its CHOP target (Figs 3D and EV2B), reflecting the activation of pro-apoptotic eIF2α/ATF4/CHOP signaling. It was also associated with the induction of ATF4 target GADD34 (Fig 3D), suggesting that dephosphorylation of p-eIF2α is induced in treated MM cells. In MM.1S cells treated with BZ for 6 h, we also observed defects in ATF6 glycosylation and partial cleavage of the protein (Fig 3E). In agreement with previous reports (Ogbechi et al, 2018), mycolactone did not induce such ATF6 cleavage. However, defective glycosylation and depletion of full-length ATF6 were observed, likely resulting from mycolactone-mediated blockade of this Type II transmembrane protein in translocation. MM1.S cells exposed to the mycolactone–BZ combination displayed the sum of the alterations induced by each drug, namely depletion, defective glycosylation, and partial cleavage of ATF6. Finally, gel electrophoresis and qPCR analyses demonstrated an enhanced XBP1 splicing in MM1.S cells treated with the mycolactone/BZ combination, compared to single drugs (Fig 3F). A significant upregulation of sXBP1 mRNA levels was also observed in KMS-11 cells exposed to the drug combination (Fig EV2B). From these data, we conclude that the mycolactone–BZ combination hyperactivates PERK and IRE1 signaling in MM cells, with potent activation of the ATF4/CHOP axis indicating a transition to terminal UPR. The ER-resident chaperone BiP is a master regulator of the UPR that is normally induced by ER stress and determines the threshold for induction of apoptosis (Hetz & Papa, 2018). It is interesting to note that BiP expression was not augmented in MM cells treated with mycolactone, alone or combined to BZ (Figs 3G and EV2B).
Click here to expand this figure.
Figure EV2. The mycolactone–BZ combination activates pro-apoptotic UPR in KMS-11 and JIM3 cells
KMS-11 and JIM3 cells were treated with mycolactone and/or BZ, or vehicle as control, for 6 h. ATF4 protein levels were assessed in cell lysates by Western blot (top panel) and quantified relative to GADPH levels (lower panel). Data are representative of two independent experiments with similar results. CHOP, sXBP1, and BiP mRNA levels were quantified by qPCR in the two cell lines treated as in (A). sXBP1 mRNA levels were normalized to total (spliced + unspliced) XBP1 mRNA level. Data are Mean RNA fold changes (2−ΔΔCT) ± SD, relative to untreated controls. N ≥ 5 (cumulative data of two independent experiments with technical duplicates and triplicates), pairwise compared by nested one-Way ANOVA with Tukey’s multiple-comparison test, exact P-values indicated. Thapsigargin (Tg, 2 µM, 6 h) was used as a positive control.Source data are available online for this figure.
The cytotoxic synergy between mycolactone and BZ is maintained in BZ-resistant MM.1S cellsThe development of resistance to PIs is a major obstacle to successful treatment of MM patients (Lawasut et al, 2012). To determine if the cytotoxic synergy between mycolactone and BZ was maintained in BZ-resistant MM cells, we generated a BZ-resistant version of the MM.1S cell line (MM.1S BzR cells), with stable and > 15× increased resistance to BZ treatment (Fig 4A). Notably, MM.1S BzR cell susceptibility to mycolactone was identical to that of MM.1S BzS (Figs 4B and 1A), showing that acquired resistance to BZ does not confer cross-resistance to mycolactone. As in MM.1S BzS cells (Fig 2C), mycolactone synergized with BZ for induction of MM.1S BzR cell death (Fig 4C and D) and engagement of apoptosis in MM.1S BzR cells exposed to the mycolactone–BZ combination correlated with activation of ER stress responses, reflected by elevated levels of ATF4, CHOP, and sXBP1 mRNAs (Fig 4E). Like MM.1S BzS cells, MM.1S BzR cells failed to upregulate BiP expression upon exposure to the mycolactone/BZ drug combination (Fig 4E). Together, these data demonstrate that mycolactone cytotoxicity and synergy with BZ override MM cell resistance to BZ. They consolidate the clinical relevance of Sec61 inhibitor-based treatments for MM.

Figure 4. The cytotoxic synergy between mycolactone and BZ is maintained in BZ-resistant cells
Parental MM.1S (BzS) and MM.1S BzR cells were incubated with increasing concentrations of BZ for 24 h. The proportion of live (Annexin V- PI-) cells was then measured by flow cytometry as in Appendix Fig S1. Data are Mean % of live cells from technical duplicates, relative to vehicle controls. Effect of mycolactone on MM.1S BzR cell viability after 48 and 72 h of treatment, as measured by exposure of Annexin V and incorporation of PI. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, gated as in Appendix Fig S1, relative to total cells. MM.1S BzR cells were treated for 24 h with mycolactone and/or BZ at the indicated concentrations, or vehicle as control. Data are Mean % ± SD of live, apoptotic, and dead cells, gated as in Appendix Fig S1, relative to total cells. N = 6 (cumulative data of three independent experiments with technical duplicates). Synergy between mycolactone and BZ in the MM.1S BzR lines, when treated as in (C). Data are Mean Loewe scores ± SD, shown as heatmaps. Differences between treated cells and controls by Student’s t-test: *P < 0.05; N = 6 (cumulative data of three independent experiments with technical duplicates). ATF4, CHOP, sXBP1, and BiP mRNA levels were quantified by qPCR in MM.1s BzR cells treated as in (C). sXBP1 mRNA levels were normalized to total (spliced + unspliced) XBP1 mRNA level. Data are Mean RNA fold changes (2−ΔΔCT) ± SD, relative to untreated controls. N = 5 (cumulative data of two independent experiments with technical duplicates and triplicates), pairwise compared by nested one-way ANOVA with Tukey’s multiple-comparison test, exact P-values indicated. Thapsigargin (Tg, 2 µM, 6 h) was used as a positive control.Data information: In A and B, shown data are representative of two independent experiments with similar results.
The synergy between mycolactone and BZ cytotoxicity extends to mouse B-cell lymphomasBesides MM, PIs show promise in the treatment of other hematological malignancies such as acute leukemia (Cloos et al, 2017) and solid malignancies (Roeten et al, 2018). We tested the proteotoxic effects of mycolactone, alone and combined with BZ, in B-cell acute lymphoblastic leukemia (B-ALL) using mouse pro-B-cell lines generated by transformation of hematopoietic cells with the murine viral form of the Abelson oncogene (v-abl). Mycolactone alone demonstrated potent ability to induce v-abl cell apoptosis in vitro (Fig 5A). Moreover, as in human MM cell lines, it synergized with BZ for induction of v-abl cell death (Fig 5B) and this correlated with activation of the ER stress-associated ATF4/CHOP apoptotic pathway (Fig 5C and D). In addition to illustrating mycolactone’s ability to efficiently block mouse Sec61, these data revealed the potential interest of Sec61 inhibitors for the treatment of B-ALL, and potentially other proteasome inhibition-susceptible malignancies.

Figure 5. The synergy between mycolactone and BZ cytotoxicity extends to mouse B-cell lymphomas
Effect of mycolactone on v-abl pro-B-cell viability after 24 and 48 h of mycolactone treatment, as measured by exposure of Annexin V and incorporation of PI. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, gated as in Appendix Fig S1, relative to total cells. v-abl pro-B cells were treated for 24 h with mycolactone and/or BZ at the indicated concentrations. The percentage of live, apoptotic, and dead cells was then measured by flow cytometry as in Appendix Fig S1. Data are Mean % from technical duplicates, relative to total cells. v-abl pro-B cells were treated with mycolactone and/or BZ at the indicated concentrations for 6 h. CHOP mRNA levels were quantified by qPCR. Data are Mean RNA fold changes (2−ΔΔCT) ± SD relative to untreated controls. N = 6 (cumulative data of three independent experiments with technical duplicates), pairwise compared by nested one-way ANOVA with Tukey’s multiple-comparison test, exact P-values indicated. v-abl pro-B cells were treated for 6 h with mycolactone and/or BZ at the indicated concentrations. ATF4 protein levels were assessed in cell lysates by Western blot (top panel) and quantified relative to GADPH levels (lower panel).Data information: Data shown in A, B, and D are representative of experiments performed with two independent v-abl cell clones, with similar results. In C and D, thapsigargin (Tg, 2 µM, 6 h) was used as a positive control.
Patient-derived MM tumors are highly susceptible to mycolactone toxicity and synergy with BZWe next assessed the anti-MM activity of mycolactone, alone and combined to BZ, in patient-derived tumors. Mononuclear cells were isolated from bone marrow aspirates of four newly diagnosed MM patients, and two patients with relapsed MM after at least one line of treatment including PI and IMiDs (Table 1). Cells were placed in culture medium within 3 h post-biopsy, then exposed to mycolactone and/or BZ for 18 h. Induction of apoptosis was determined in MM cells, gated as CD38+ CD138+/− plasma cells, following Annexin V/PI staining (Appendix Fig S2). Figure 6A shows the results as % of live MM cells and Fig EV3A as % of live/apoptotic/dead cells, for easier comparison with treatment effect on non-cancerous lymphoid cells (Fig EV3B). MM cells from the six studied patients varied in susceptibility to BZ treatment, irrespective of their newly diagnosed or relapsed status (Figs 6A and EV3A). Tumor sensitivity to mycolactone was also variable, and not systematically associated with sensitivity to BZ (Figs 6A and EV3A). In all studied patients, significant cell death was achieved with 18 h exposure to ≥ 12 nM mycolactone, an anti-MM activity globally equivalent to that of 10 nM BZ. Strikingly, mycolactone synergized with BZ to kill MM cells from all patients irrespective of their treatment naive or relapsed status and relative resistance to single drugs (Fig 6B), further demonstrating that mycolactone-mediated cytotoxicity operates irrespective of MM cell resistance to BZ.
Table 1. Characteristics of the MM patients included in this study. Patient P2 P5 P7 P9 P12 P13 Age (years) 57 72 72 67 72 66 Gender (F/M) F F F F M F Years since diagnosis (years) – – 10 – 5 – Lines of treatment – – 2 – 1 – Treatments IMiDs – – 2 – 1 – PI – – 2 – 1 – Alkylating agent – – 1 – 1 – HSCTa – – 1 – – –
Figure 6. MM primary tumors are highly susceptible to mycolactone toxicity and synergy with BZ
Mononuclear cells from bone marrow aspirates of newly diagnosed (#2, #5, #9, and #13) or relapsed (#7 and #12) MM patients were treated with mycolactone and/or BZ at the indicated concentrations, or vehicle as control, for 18 h. Then, MM cells were identified by staining with anti-CD38 and anti-CD138 antibodies using the gating strategy depicted in Appendix Fig S2.
Following treatment, induction of apoptosis/necrosis was measured by exposure of annexin V and PI incorporation. Data are Mean % of live cells from technical duplicates, relative to controls. Synergy between mycolactone and BZ in treated tumors. Data are Mean Loewe scores from technical duplicates, shown as heatmaps. Click here to expand this figure.
Figure EV3. Differential toxicity of mycolactone–BZ combination in MM cells and non-cancerous lymphoid cells in bone marrow aspirates
A, B. Incidences of live, apoptotic, and dead cells within the MM cell (A) and the non-cancerous lymphoid cell (B) subsets are compared in newly diagnosed (#2, #5, #9, and #13) or relapsed (#7 and #12) MM patients after an 18 h treatment with mycolactone and/or BZ. MM cells (A) and non-cancerous lymphoid cells (B) were identified with the gating strategy depicted in Appendix Fig S2. Data are Mean % of live, apoptotic, and dead cells from technical duplicates, relative to total cells.Importantly, the cytotoxicity of the mycolactone–BZ combination selectively affected MM cells, as minimal cell death was recorded in CD38- CD138- cells from the same bone marrow aspirates (Fig EV3A and B). It is noteworthy that bone marrow aspirates from the two relapsed patients contained a relatively higher incidence of monocytes/macrophages, as characterized by their SSC/FSC profile (as shown for Patient #7 in Appendix Fig S2). Although drugs alone had little effect, the mycolactone–BZ combination displayed some toxicity on this cell subset at the highest tested concentrations (Fig EV4A). To further assess the impact of mycolactone–BZ combination on immune cell viability, we subjected peripheral blood mononuclear cells (PBMCs) to the same drug treatments as patient-derived tumors (Fig 6 Appendix Fig S3). Minimal or no viability loss was observed in total PBMCs, nor in gated T cells, B cells, natural killer cells, dendritic cells, and monocytes/macrophages exposed to mycolactone, BZ, or both drugs (Fig EV4B), indicating that the cytotoxicity of the mycolactone–BZ combination is selective of MM cells.
Click here to expand this figure.
Figure EV4. Toxicity of the mycolactone–BZ combination in PBMCs and bone marrow monocytes/macrophages
The proportion of live (Annexin V- PI-) cells within the monocyte/macrophage subset of mononuclear cells, after a 18 h treatment with mycolactone and/or BZ is shown for the two relapsed patients. Mononuclear cells from bone marrow aspirates were treated as in Fig 6. The monocyte/macrophage subset was identified using the gating strategy depicted in Appendix Fig S2. Data are Mean % of live cells, relative to vehicle controls. PBMCs from healthy donors were treated with mycolactone and BZ, alone or in combinations, at the indicated concentration for 18 h. Cells were then labelled with fluorophore-conjugated anti-CD3, anti-CD19, and anti-CD11c. Total cells, B cells, T cells, dendritic cells, NK cells, and monocytes/macrophages were identified by flow cytometry analysis, using the strategy depicted in Appendix Fig S3. Data are Mean % ± SD of live cells, relative to vehicle controls. N = 6, cumulative data from three donors with technical duplicates. Combining mycolactone with BZ delays MM tumor growth in vivoTo evaluate the therapeutic efficacy of the mycolactone–BZ combination in vivo, we first analyzed its toxicity in mice. C57BL/6 mice were treated twice a week by intraperitoneal injection of BZ (0.5 mg/kg) alone and/or mycolactone (0.3 mg/kg) for 3 weeks, BZ and mycolactone treatments previously shown to induce anti-MM activity (LeBlanc et al, 2002) and anti-inflammatory effects in injected mice (Guenin-Mace et al, 2015), respectively. No sign of distress or weight loss could be detected in mice receiving single drugs or the drug combination, and their blood cell counts remained unaltered throughout the experiment (Fig 7A), illustrating the safety of these treatment regimen. We next compared the anti-MM activity of mono- and bi-therapies in immunodeficient NOD/SCID/IL2rγnull (NSG) mice. Mice were subcutaneously engrafted with MM.1S cells, and 7 days later they were assigned to four treatment groups receiving (i) 0.5 mg/kg BZ, (ii) 0.3 mg/kg mycolactone, (iii) both drugs at these concentrations, or (iv) vehicle, twice weekly via the intraperitoneal route. Under these conditions, BZ and mycolactone both induced a minor yet significant delay in tumor growth, compared to vehicle controls (Fig 7B). Notably, the mycolactone–BZ combination was superior to single drug treatments in slowing down MM development. In addition to revealing a therapeutic window for Sec61 blockade in MM, these data confirmed the interest of combining inhibitors of translocon and proteasome in MM therapy.
留言 (0)