
記住我








The Food and Agriculture Organization of the United Nations and the World Health Organization have defined probiotics as live microorganisms, which, when administered in adequate amounts, confer a health benefit on the host.1 The International Scientific Association for Probiotics and Prebiotics consensus statement subsequently clarified the scope to specify microbial species that have been shown in properly controlled studies to confer benefits to health.2 The definition of a prebiotic, as a substrate that is selectively utilized by host microorganisms conferring a health benefit, was expanded to include non-carbohydrate substances, applications to body sites other than the gastrointestinal tract, and diverse categories other than food, while the requirement for selective microbiota-mediated mechanisms was retained.3 The term paraprobiotic was introduced to encompass the possibility of using non-viable microbial cells or crude microbial cell fractions as health-promoting agents to modulate host immune responses.4, 5
Non-viable microorganisms, microbial compounds, and growth promoting substrates extend the scope of probiotic concepts.5 For example, heat-killed and tyndallized probiotic lactic acid bacteria and bifidobacteria were shown to have key probiotic effects in gastrointestinal and other diseases.6 Consumption of lactic acid bacteria fermentates or powdered preparations, derived from a fermented product, e.g., fermenting microorganisms, components of these microorganisms, culture supernatants, fermented substrates, or bioactive components, was found to have comparable activity to the live forms.7
Until fairly recently, microbiome and probiotic research studies have mainly concerned gastrointestinal diseases. However, there is increasing evidence that benefits of probiotics extend beyond the digestive system.
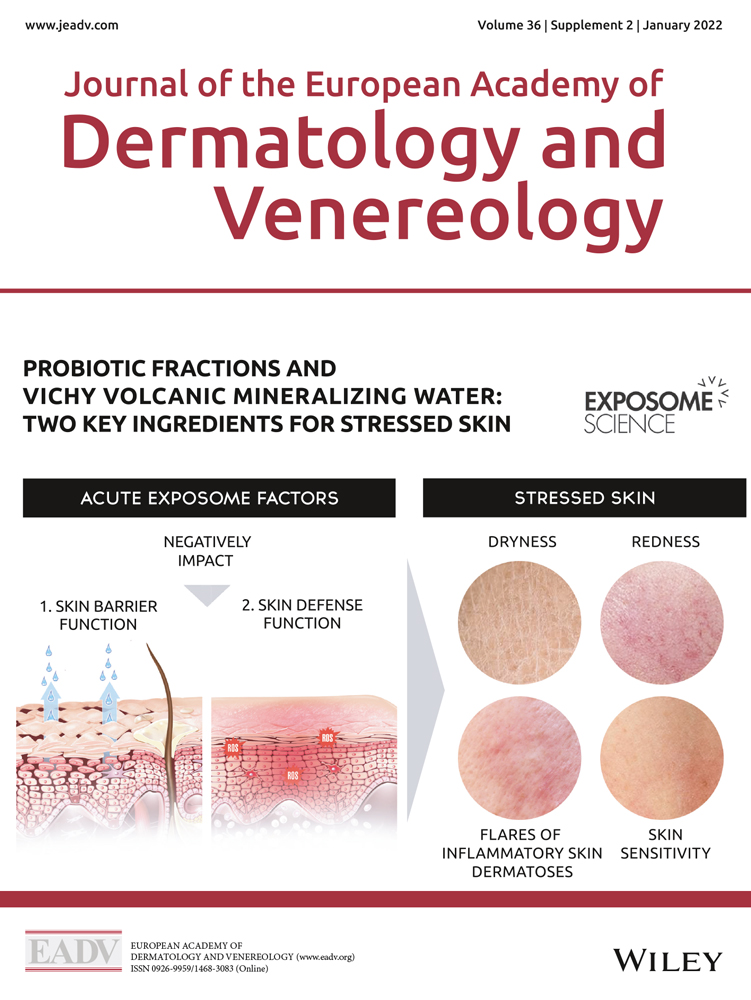
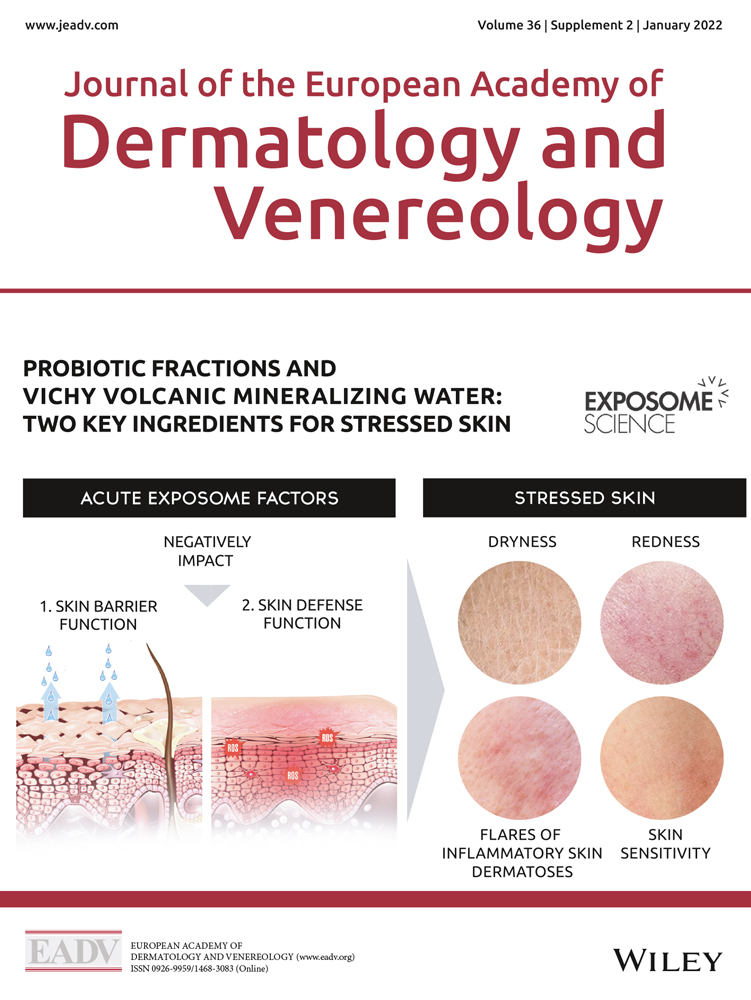
The skin is a diverse barrier organ and the first line of defense against aggression from environmental stressors, e.g., meteorological factors, ultraviolet radiation (UVR), pollution or tobacco smoke, as well as internal stressors, e.g., an unhealthy diet, hormonal variations, lack of sleep, and psychosocial stress. These exposome factors can affect several functions of the skin.8, 9
The gut−skin axis and the effects of the microbiome on the skin are becoming better understood, and there is a growing interest in using oral10, 11 and topical11-13 probiotics to improve skincare and various skin conditions. Probiotics strengthen the barrier function (and/or protect the commensal flora) and promote skin immune defenses against pathogens (at both microbiota and skin cell levels) and aggressions.
Benefits of oral probiotics in dermatologyPrebiotics, probiotics, or both are used in allergy prevention under certain conditions.14 In particular, the World Allergy Organization guideline for allergic disease prevention by probiotics determined that there is a likely net benefit from using probiotics to prevent eczema.15
Oral probiotics have a positive effect on skin health, via the gut and the systemic route, by modulating the immune system and providing benefits.16, 17 Oral probiotics can be used as a complementary therapy or can be used alone to improve the clinical outcome of certain dermatological diseases.18
In addition to the ability to modulate both intestinal mucosal and systemic immune functions, the probiotic strain Lactobacillus paracasei NCC2461 (ST11) may confer benefits at the skin level to reinforce the skin barrier function and modulate the skin defenses by reducing neurogenic inflammation, resulting in decreased skin sensitivity.19-21 In a randomized, double-blind, clinical study after capsaicin-induced irritation, oral supplementation with L. paracasei NCC 2461 (ST11) was shown to decrease skin sensitivity and increase the rate of barrier function recovery improving sensitive skin.20 The probiotic Lactobacillus johnsonii La1 has been shown to protect the skin defenses by maintaining the number and function of Langerhans cells (LC) after UV exposure to the skin, as well as regulating skin inflammation.22, 23 Supplementation with oral probiotic L. johnsonii in vivo protected skin immune homeostasis by decreasing UV-induced immunosuppression after UV exposure in a double blind, randomized, placebo controlled clinical trial.22, 24 In addition, oral probiotics have been shown to prevent and reduce the severity of atopic dermatitis,17 acne vulgaris,25 dandruff,26 dry skin, prevent signs of photoaging, and facilitate wound healing.27
Benefits of topical probiotic extracts in dermatologyHealth benefits on the skin can be achieved by topical application of inactivated probiotics.12, 16, 28 The majority of topical bacterial extracts are ferments, ferment lysates, or ferment lysate filtrates generated after cultivation and harvest of a probiotic microorganism. Some bacterial extracts (Bacillus coagulans, L. johnsonii, Lactobacillus casei, Lactobacillus plantarum, and Lactobacillus acidophilus) have antimicrobial properties that may support skin healing.29, 30 Also, topical probiotics may have beneficial properties on reactive skin, providing photoprotection, and reducing signs of skin aging.31-33
A systematic review found topical inactivated probiotic fractions have demonstrated efficacy in clinical trials in acne, atopic dermatitis, and rosacea but reported a paucity of information on both mechanistic and clinical investigation of topical probiotics fractions.13, 34
Tyndallized L. acidophilus extract regulates skin health through antimicrobial peptides (AMP) and has been shown in vitro to scavenge reactive oxygen species following UVB-induced oxidative stress leading to antiaging effects in keratinocytes.35
The lysates of various inactivated probiotic lactic acid bacteria were shown to stimulate the proliferation of keratinocytes. Lactobacillus plantarum and Lactobacillus salivarius lysates accelerated re‑epithelisation by inducing keratinocyte migration.36 Microfluidized lysates of Lactobacillus rhamnosus may improve the skin barrier function in a reconstructed human epidermis.37 Application of the L. rhamnosus lysate on skin increased the expression of tight junction proteins (claudin 1 and occludin) and skin barrier proteins [loricrin and filaggrin (FLG)], while pretreatment with the lysate alleviated effects of the skin irritant, sodium lauryl sulfate.37 Topical application of the Lactobacillus bulgaricus, L. acidophilus, or L. plantarum extract improved acne by reducing skin colonization by Cutibacterium acnes.38, 39
A randomized controlled trial in patients with atopic dermatitis have demonstrated that topical application of Vitreoscilla filiformis extract (Vfe) alleviated skin inflammation and improved atopic dermatitis.40 Additionally, Vfe ointments improved symptoms of seborrheic dermatitis.41
Vitreoscilla filiformis extractVitreoscilla filiformis extract is considered as a probiotic fraction since topical application of this extract has shown activity to maintain good homeostasis of skin defenses. Mahe et al.42 evaluated whether biomass obtained from biotechnological processing of V. filiformis bacteria could stimulate endogenous anti-oxidant defenses. Mitochondrial manganese superoxide dismutase-2 activity, an inducible free-radical scavenger in skin cells, was assayed in vitro in human skin fibroblasts both at the mRNA and protein levels. Vfe was shown to modulate mitochondrial superoxide dismutase. Furthermore, in a clinical study, Vfe showed beneficial properties, compared to an emollient containing sunscreen alone, to protect against the formation sunburn cells in the epidermis following UVB irradiation.42
Innate sensing of Vfe predominantly induced IL-10 producing dendritic cells, which was dependent on TLR2 activation.43 Dendritic cells exposed to Vfe caused induction of IL-10 + T helper cells to become regulatory Tr1 (type 1 regulatory T) cells that suppress T effector cells and reduce inflammation.43
Production of VfeVitreoscilla filiformis (ATCC 15551) is a non-pathogenic, non-fruiting, non-photosynthetic filamentous gliding bacterium belonging to the Beggiatoales order.44
Vitreoscilla filiformis is cultured on an industrial scale in a tightly controlled biotechnological fermentation process designed for delicate bacteria to ensure reproducibility, stability, and biological activity. After harvesting, the cells are lysed to produce the bacterial extract (lysate including membrane and cytosol). Vfe is very rich in 16 amino acids, including nine essential amino acids.
Vfe cultured with Vichy volcanic mineralizing waterVichy volcanic mineralizing water (VVMW) originates in the volcanic Auvergne region of France and is a unique, supercharged, pure water, highly enriched in 15 minerals recognized for its regenerating, antioxidant and soothing properties.45 It is used as a dermocosmetic ingredient to strengthen the skin against exposome aggressions.45
Vichy volcanic mineralizing water has unique properties to repair and regenerate the skin physical barrier and strengthen antioxidant defenses, as well as anti-inflammatory properties, to help combat exposome aggressions, as described in another article in this supplement.
The extract of V. filiformis used in the studies described below was grown at pH 7.2 and 26°C in a sterile complex, well-defined, culture medium containing 15 nutrients (sugar, plant extracts, yeast, minerals, and vitamins) and reverse osmosis water containing VVMW with no preservatives. The term we have used to specify this extract of V. filiformis grown in a medium containing VVMW is VfeV.
The aim of the studies described in this article was to evaluate the effect on skin functions of VfeV in combination with VVMW to help understand how this combination of ingredients can counteract exposome aggressions to the skin.
A combination of VfeV and VVMW reinforces the physical skin barrierOne of the most important functions of the skin is to act as a physical barrier to protect the organism and prevent uncontrolled water loss.
Tight junctions and keratinocyte differentiationThe tight junctions form a part of the epithelial permeability barrier between the normal human epidermal keratinocyte cells. These structures are important components of the skin barrier function, linking structural integrity to proliferation and inflammatory responses of the skin.
We evaluated in vitro the effect of a combination of VfeV and VVMW on skin barrier markers (see Figs 1 and 2). In an in vitro assay, normal human epidermal keratinocytes (NHEK) isolated from foreskin were seeded in 96-well plates and cultured at 37°C for 24 h (192 h for FLG) in serum-free medium (supplemented with 25 µg/mL gentamycin, 25 µg/mL pituitary extract, 0.25 ng/mL epidermal growth factor). The keratinocytes were then transferred to medium in the absence (control) or presence of 10% VVMW, 0.002% VfeV or both. NHEK were also treated with the reference 1.5 mmol/L calcium chloride, a well-known epidermal differentiation factor, as a positive control. After 72 h incubation, fixed cells were immunolabeled with antibodies directed against epidermal terminal differentiation markers (transglutaminase (TGM1), FLG, and involucrin (INV)] and tight junction protein synthesis [claudin-1 (CLDN1) and zonula occludens-1 (ZO-1)]. The antibodies were revealed by fluorescent secondary antibodies and labeling was quantified by measuring fluorescence intensity of proteins in relation to the number of cell nuclei identified using Hoechst staining (bis-benzimide).

Effect of Vitreoscilla filiformis extract combined with Vichy volcanic mineralizing water on keratinocyte differentiation markers filaggrin and involucrin, as assayed in vitro in normal human epidermal keratinocytes (a) with representative high-resolution images of Hoechst staining (b). Normal human epidermal keratinocytes (NHEK, Bioalternatives, Gençay, France) isolated from foreskin were seeded in 96-well plates and cultured in Keratinocyte SFM medium (GIBCO, Thermo Fisher Scientific, Asnières-sur-Seine, France) supplemented with epidermal growth factor (EGF) 0.25 ng/mL, pituitary extract 25 μg/mL, gentamycin 25 μg/mL (GIBCO) for 24 h (involucrin), or 192 h (filaggrin). The medium was then replaced with MCDB153 medium (GIBCO) supplemented with gentamycin 25 μg/mL (GIBCO) with or without (control) the test compounds, their combination, or the reference (CaCl2, 1.5 mmol/L) and the cells were incubated for 144 h (involucrin) or 72 h (filaggrin). After incubation, the cells were rinsed, fixed, and permeabilized, before labeling with the primary antibody (anti-filaggrin, ref. sc-66192, Santa Cruz Biotechnology, Inc, Heidelberg, Germany; anti-involucrin, ref. J64013, Alfa Aesar, Thermo Fisher GmbH, Kandel, Germany). This antibody was then revealed by a secondary antibody coupled to a fluorochrome (GAR-Alexa 488, ref. A11008, IInvitrogen, Illkirch France). In parallel, cell nuclei were stained with Hoechst 33258 (bisbenzimide, ref. B1155, Sigma, Darmstadt, Allemagne). For each well, five scanned images were captured using a high-resolution imaging system (INCell Analyzer, GE Healthcare, Buc, France). Labeling was quantified by measuring the fluorescence intensity of the proteins relative to the number of cells identified by Hoechst with digital data integration. All experimental conditions were performed in triplicate. VfeV, Vitreoscilla filiformis extract grown in a medium containing VVMW; VVMW, Vichy volcanic mineralizing water; *P = 0.01 to 0.05, **P = 0.001 to 0.01, ***P < 0.001.
 Effect of Vitreoscilla filiformis extract combined with Vichy volcanic mineralizing water on tight junction proteins zonula occludens-1 and claudin-1 in normal human epidermal keratinocytes (a) with representative high-resolution images of Hoechst staining (b). The experimental conditions were as described in Fig. 1 for involucrin, except the primary antibodies were anti-claudin 1 (ref. sc-81796; Santa Cruz Biotechnology, Inc, Heidelberg, Germany) and anti-ZO-1 (ref. 33-9100; Invitrogen). VfeV Vitreoscilla filiformis extract grown in a medium containing VVMW; VVMW, Vichy volcanic mineralizing water; *P = 0.01 to 0.05, **P = 0.001 to 0.01, ***P
Effect of Vitreoscilla filiformis extract combined with Vichy volcanic mineralizing water on tight junction proteins zonula occludens-1 and claudin-1 in normal human epidermal keratinocytes (a) with representative high-resolution images of Hoechst staining (b). The experimental conditions were as described in Fig. 1 for involucrin, except the primary antibodies were anti-claudin 1 (ref. sc-81796; Santa Cruz Biotechnology, Inc, Heidelberg, Germany) and anti-ZO-1 (ref. 33-9100; Invitrogen). VfeV Vitreoscilla filiformis extract grown in a medium containing VVMW; VVMW, Vichy volcanic mineralizing water; *P = 0.01 to 0.05, **P = 0.001 to 0.01, ***P
The results in NHEK showed a statistically significant increase in TGM1 synthesis with 10% VVMW, a not statistically significant increase by VfeV (0.002%), while the combination of 10% VVMW plus 0.002% VfeV increased the synthesis of TGM1 by 142% compared to the control (P < 0.05). For FLG synthesis, VVMW (10%) statistically significantly increased FLG (348% compared to the control, P < 0.001), stimulation by VfeV (0.002%) was not significant, while the combination of 10% VVMW plus 0.002% VfeV had a statistically significant additive effect on the expression of FLG (585% compared to the control; P < 0.01; Fig. 1a,b). For INV synthesis, a statistically significant increase of 158% was observed with 10% VVMW plus 0.002% VfeV, in comparison with the control (P < 0.05; Fig. 1a,b).
In NHEK, 10% VVMW in combination with 0.002% VfeV significantly increased CLDN1 and ZO-1 by 264% and 134%, respectively, in comparison with the controls (all P < 0.05) (Fig. 2a,b). Synthesis of CLDN1 was significantly stimulated by 0.002% VfeV and slightly stimulated by 10% VVMW, while the combination of 0.002% VfeV plus 10% VVMW demonstrated a statistically significant additive effect (Fig. 2a,b).
These results demonstrate that the combination of 10% VVMW plus 0.002% VfeV reinforced the skin barrier physical defenses with significant additive effects on FLG expression, which is a terminal differentiation protein, as well as on CLDN1 synthesis, which is a major component of epidermal tight junctions that creates a dynamic paracellular barrier in the epidermis.
A combination of VfeV and VVMW stimulates antimicrobial peptide defensesThe epidermis plays an important role in innate immune responses. Innate immune responses protect the body against microbial invasion by killing pathogenic microorganisms through the production of cationic AMPs, such as human β-defensins and cathelicidin.46, 47
Gene expression in keratinocytesThe skin is an active immune organ playing a crucial role in innate immune responses that protect the body against microbial invasion by killing pathogenic microorganisms through among others the production of cationic AMP, such as human β-defensins and cathelicidin.
Gene expression in NHEK was assayed using RT-qPCR technology on total RNA extracted from the cell monolayers and targeted five genes coding for AMPs [defensin (DEFB1), β4A human defensin 2 (DEFB4A), CAMP, RNASE7, S100A7]. To overcome variability, relative expression of above 200% as a percentage of the control was considered as stimulation. In the presence of 0.3% VfeV, the AMPs RNASE7, S100A7, and DEFB4A were upregulated by 246%, 589%, and 1490%, respectively, in comparison with the controls (all P < 0.05). Although there was no effect was measured with VVMW alone, in the presence of 50% VVMW in combination with 0.03% VfeV, the expression of DEFB4A was increased by 3266% compared to the control (P < 0.05), indicating an additive effect on stimulating this AMP.
A combination of VfeV and VVMW stimulates cellular immune defensesThe immunoregulatory and anti-inflammatory effects of a combination of VfeV and VVMW were evaluated in vitro in peripheral blood mononuclear cells (PBMCs) and LCs.
Cytokine release by peripheral blood mononuclear cellsTo evaluate immunoregulatory effects, activation of PBMCs was analyzed under LPS-stimulated conditions. PBMC were isolated from total peripheral blood and cultured at 37 ˚C in culture medium reconstituted with VVMW for 1 h. After 1-h preincubation in medium containing the test compounds, inflammation was induced by the addition of 1µg/mL lipopolysaccharide (LPS) and the cells were incubated for 24 h. Cytokine/chemokine release were quantified in the culture supernatants. In LPS-stimulated PBMCs, a combination of 16.7% VVMW and 0.1% VfeV down-regulated IL-8, TNF-α, IL-12/IL-23p40, and increased IL10 (568% compared to control, P < 0.001), and the IL-10/IL-12 ratio (3297% compared to control, P < 0.05). The high ratio of IL-10/inflammatory cytokines indicate that the combination of VfeV and VVMW has significant immunomodulatory properties.
Protection of Langerhans cells against ultraviolet-induced damageSupporting the inflammatory defense of the epidermis are the dendritic cells residing in the epidermis in the form of LCs as antigen-presenting cells, which play a crucial role in the immune response. Exposure to ultraviolet (UV) including UVA and UVB induces a decrease in the number and modifies the morphology of LCs, as well as an alteration in their capacity to present antigens, which reduces the ability of the immune system to respond to contact allergens.
Protective effects of a combination of VfeV and VVMW were assessed ex vivo using a human skin explant model by evaluating the number of LCs using in situ immunolabeling (flurorescence) of CD1a cell membrane specific marker (see Fig. 3). Test skin explants were cut into pieces (2 × 2 cm), placed in culture medium and untreated (control) or topically pretreated with the test compounds, combination (at 20 or 5 mg/cm2). After 24 h preincubation, the treatment was renewed and incubated for 45 min. Immediately after UVB/UVA exposition, the treatment was renewed, and skin explants were incubated for 24 h before immunolabeling and image analysis. The CD1a-positive cells (LC, green fluorescence) were counted and normalized.

Effect of Vitreoscilla filiformis extract combined with Vichy volcanic mineralizing water on Langerhans cell number, as measured by CD1a expression in UV-irradiated skin explants (a) and representative images of CD1a-positive cells (Langerhans cells, green fluorescence) (b). Upon receipt of the biopsy from abdominal plastic surgery, skin explants were cut into pieces (2 × 2 cm), placed in DMEM/F12 medium and topically pretreated with VVMW, VfeV, or the combination, or the reference (solar cream SPF 30 at 5 mg/cm2) or left untreated (control). After preincubation of the skin explants for 24 h, the treatment was renewed, and incubated for a further 45 min. The skin explants were then irradiated by 1.25 J/cm² UVB (and 3.8 J/cm² UVA) using a UV-irradiation chamber BS-02 lamp (Opsytec Dr. Gröbel). In parallel, a non-exposed control was kept in the dark. Immediately after UV exposition, the treatment was renewed, and the skin explants were incubated for 24 h. After incubation, the supernatants were discarded, the skin explants were rinsed, and punches were performed on each skin explant (punch of 8 mm for immunolabeling). The frozen sections were incubated with the appropriate primary antibody (anti-CD1a, BD Biosciences, Le Pont de Claix, France, ref. 555805), and secondary antibody (GAM Alexa Fluor 488, Invitrogen, ref. A11001) and the cell nuclei were stained with propidium iodide (Sigma, ref. P4170) solution. The sections were observed using a Nikon Eclipse Ci microscope and digital images were captured using a Nikon (Grenchen, Switzerland) DS-Ri2 camera and processed using NIS-Element (Grenchen, Switzerland) 4.20.00 software. The CD1a-positive cells (Langerhans cells, green fluorescence) were counted and normalized to the total epidermal surface. All experimental conditions were performed in triplicate. VfeV, Vitreoscilla filiformis extract grown in a medium containing VVMW; VVMW, Vichy volcanic mineralizing water; *P = 0.01 to 0.05, **P = 0.001 to 0.01.
The mean number of CD1a-positive nuclei per μm² for the non-exposed control was 199 and for the exposed was 37 number of CD1a-positive nuclei/μm². The protection with 79% VVMW alone, 5% VfeV alone, and 79% VVMW plus 5% VfeV was better than bare skin and the percentage improvements compared to the irradiated control were 198%, 173%, and 257%, respectively (Fig. 3a); representative images of immunolabeling of LC are shown in Fig. 3b. These results showing that VfeV plus VVMW protected LCs in skin explants exposed to UV illustrate the additive capacity to preserve skin immune homeostasis even when exposed to solar radiation.
Conclusions and perspectives These in vitro studies provide mechanistic insight into the beneficial properties of a combination of VfeV and VVMW and show that the combination had additional beneficial effects than either component alone, on: Strengthening the skin barrier. Increased tight junctions. Increased keratinocyte differentiation. Protecting biochemical immune defense Stimulated AMP (defensin) synthesis. Protecting cellular immune defense Increased the IL-10/IL-12 ratio. Protected Langerhans cells challenged by UVR.The combination had additive effects on stimulating the filaggrin differentiation marker, claudin-1 tight junction protein, β-defensin-4A AMP, as well as protecting LCs challenged by UVR. A dermocosmetic formulation containing a combination VVMW and Vfe probiotic fractions may thus be highly effective in boosting immune defenses, as well as restoring the skin barrier, to strengthen the skin against exposome aggressions and help prevent photoaging and repair stressed skin.
AcknowledgementsWriting and editorial assistance was provided by Helen Simpson, PhD, ISMPP CMPP™, of My Word Medical Writing and funded by Vichy Laboratories.
Ethical standardsAll clinical studies were conducted according to ICH CPMP Good Clinical Practices guidelines and the Declaration of Helsinki (1964, and its subsequent modifications), and all subjects were healthy volunteers who gave written informed consent. Independent Ethics Committee approval was not required for these cosmetic product studies (the investigational product did not contain and was not derived from material with specific risks, and all the procedures were noninvasive).
Data availability statementData may be shared upon submission of a research proposal to the corresponding author. The research proposal will be reviewed, and if agreed, the requestors will need to sign a data sharing agreement.
Data availability statementData may be shared upon submission of a research proposal to the corresponding author. The research proposal will be reviewed, and if agreed, the requestors will need to sign a data sharing agreement.
References
1 FAO/WHO. Food and Agriculture Organization of the United Nations/World Health Organization. Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria. FAO/WHO; 2001. http://www.fao.org/publications/card/en/c/7c102d95-2fd5-5b22-8faf-f0b2e68dfbb6/ (last accessed 16 November 2021). 2Hill C, Guarner F, Reid G et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol 2014; 11: 506– 514. 3Gibson GR, Hutkins R, Sanders ME et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol 2017; 14: 491– 502. 4Taverniti V, Guglielmetti S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: proposal of paraprobiotic concept). Genes Nutr 2011; 6: 261– 274. 5Martín R, Langella P. Emerging health concepts in the probiotics field: streamlining the definitions. Front Microbiol 2019; 10: 1047. 6Piqué N, Berlanga M, Miñana-Galbis D. Health benefits of heat-killed (Tyndallized) probiotics: an overview. Int J Mol Sci 2019; 20: 2534. 7Mathur H, Beresford TP, Cotter PD. Health benefits of lactic acid bacteria (LAB) fermentates. Nutrients 2020; 12: 1679. 8Krutmann J, Bouloc A, Sore G, Bernard BA, Passeron T. The skin aging exposome. J Dermatol Sci 2017; 85: 152– 161. 9Passeron T, Krutmann J, Andersen ML, Katta R, Zouboulis CC. Clinical and biological impact of the exposome on the skin. J Eur Acad Dermatol Venereol 2020; 34(Suppl 4): 4– 25. 10Bustamante M, Oomah BD, Oliveira WP, Burgos-Díaz C, Rubilar M, Shene C. Probiotics and prebiotics potential for the care of skin, female urogenital tract, and respiratory tract. Folia Microbiol (Praha) 2020; 65: 245– 264. 11Yu Y, Dunaway S, Champer J, Kim J, Alikhan A. Changing our microbiome: probiotics in dermatology. Br J Dermatol 2020; 182: 39– 46. 12Franca K. Topical probiotics in dermatological therapy and skincare: a concise review. Dermatol Ther (Heidelb) 2021; 11: 71– 77. 13Lee GR, Maarouf M, Hendricks AJ, Lee DE, Shi VY. Topical probiotics: the unknowns behind their rising popularity. Dermatol Online J 2019; 25: 13030. 14West CE, Dzidic M, Prescott SL, Jenmalm MC. Bugging allergy; role of pre-, pro- and synbiotics in allergy prevention. Allergol Int 2017; 66: 529– 538. 15Fiocchi A, Pawankar R, Cuello-Garcia C et al. World allergy organization-mcmaster university guidelines for allergic disease prevention (GLAD-P): probiotics. World Allergy Organ J 2015; 8: 4. 16Gueniche A, Benyacoub J, Blum S, Breton L, Castiel I. Probiotics for skin benefits. In A Tabor, RM Blair, eds. Nutritional Cosmetics, William Andrew Publishing, Boston, MA, 2009: 421– 439. 17Li L, Han Z, Niu X et al. Probiotic supplementation for prevention of atopic dermatitis in infants and children: a systematic review and meta-analysis. Am J Clin Dermatol 2019; 20: 367– 377. 18Kalil C, Chaves C, Stramari de Vargas A, Barreto CV. Use of probiotics in dermatology - review. Surg Cosmet Dermatol Rio De Janeiro 2020; 12: 203– 207. 19Benyacoub J, Bosco N, Blanchard C et al. Immune modulation property of Lactobacillus paracasei NCC2461 (ST11) strain and impact on skin defences. Benef Microbes 2014; 5: 129– 136. 20Gueniche A, Philippe D, Bastien P et al. Randomised double-blind placebo-controlled study of the effect of Lactobacillus paracasei NCC 2461 on skin reactivity. Benef Microbes 2014; 5: 137– 145. 21Gueniche A, Benyacoub J, Philippe D et al. Lactobacillus paracasei CNCM I-2116 (ST11) inhibits substance P-induced skin inflammation and accelerates skin barrier function recovery in vitro. Eur J Dermatol 2010; 20: 731– 737. 22Guéniche A, Philippe D, Bastien P, Blum S, Buyukpamukcu E, Castiel-Higounenc I. Probiotics for photoprotection. Dermatoendocrinol 2009; 1: 275– 279. 23Bouilly-Gauthier D, Jeannes C, Maubert Y et al. Clinical evidence of benefits of a dietary supplement containing probiotic and carotenoids on ultraviolet-induced skin damage. Br J Dermatol 2010; 163: 536– 543. 24Peguet-Navarro J, Dezutter-Dambuyant C, Buetler T et al. Supplementation with oral probiotic bacteria protects human cutaneous immune homeostasis after UV exposure-double blind, randomized, placebo controlled clinical trial. Eur J Dermatol 2008; 18: 504– 511. 25Kober M-M, Bowe WP. The effect of probiotics on immune regulation, acne, and photoaging. Int J Womens Dermatol 2015; 1: 85– 89. 26Reygagne P, Bastien P, Couavoux MP et al. The positive benefit of Lactobacillus paracasei NCC2461 ST11 in healthy volunteers with moderate to severe dandruff. Benef Microbes 2017; 8: 671– 680. 27Han N, Jia L, Guo L et al. Balanced oral pathogenic bacteria and probiotics promoted wound healing via maintaining mesenchymal stem cell homeostasis. Stem Cell Res Ther 2020; 11: 61. 28Al-Ghazzewi FH, Tester RF. Impact of prebiotics and probiotics on skin health. Benef Microbes 2014; 5: 99– 107. 29Ouwehand AC, Båtsman A, Salminen S. Probiotics for the skin: a new area of potential application? Lett Appl Microbiol 2003; 36: 327– 331. 30Lopes EG, Moreira DA, Gullón P, Gullón B, Cardelle-Cobas A, Tavaria FK. Topical application of probiotics in skin: adhesion, antimicrobial and antibiofilm in vitro assays. J Appl Microbiol 2017; 122: 450– 461. 31Guéniche A, Bastien P, Ovigne JM et al. Bifidobacterium longum lysate, a new ingredient for reactive skin. Exp Dermatol 2010; 19: e1– 8. 32Patra V, Gallais Serezal I, Wolf P. Potential of skin microbiome, pro- and/or pre-biotics to affect local cutaneous responses to UV exposure. Nutrients 2020; 12: 1795. 33Krutmann J. Pre- and probiotics for human skin. Clin Plast Surg 2012; 39: 59– 64. 34Kwon HH, Yoon JY, Park SY, Min S, Suh DH. Comparison of clinical and histological effects between lactobacillus-fermented Chamaecyparis obtusa and tea tree oil for the treatment of acne: an eight-week double-blind randomized controlled split-face study. Dermatology 2014; 229: 102– 109. 35Lim HY, Jeong D, Park SH et al. Antiwrinkle and antimelanogenesis effects of Tyndallized Lactobacillus acidophilus KCCM12625P. Int J Mol Sci 2020; 21: 1620. 36Brandi J, Cheri S, Manfredi M et al. Exploring the wound healing, anti-inflammatory, anti-pathogenic and proteomic effects of lactic acid bacteria on keratinocytes. Sci Rep 2020; 10: 11572. 37Jung YO, Jeong H, Cho Y et al. Lysates of a probiotic, Lactobacillus rhamnosus, can improve skin barrier function in a reconstructed human epidermis model. Int J Mol Sci 2019; 20: 4289. 38Bowe WP, Logan AC. Acne vulgaris, probiotics and the gut-brain-skin axis - back to the future? Gut Pathog 2011;
留言 (0)