
記住我







The adult human sacrum typically consists of a total of five fused sacral vertebrae. Developmentally, multiple primary and secondary ossification centers within each vertebra ossify from birth until age 25, while intersegmental fusion is completed at around age 18 (Broome et al., 1998; Cardoso et al., 2014). However, the sacrum shows a high frequency of numeric deviations, commonly attributed to segmentation anomalies (Bertolotti, 1917; Nastoulis et al., 2019; Paterson, 1892; Williams et al., 2019). The segmentation of the vertebral column occurs during embryonic development when the paraxial mesoderm starts to form transversal clefts around Day 21 that separate it into a total of 42–44 somites (Töndury & Theiler, 1990). This eventually results in five series of morphologically similar vertebrae (7 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 3–5 coccygeal vertebrae) that collectively form the human spine. A segmentation anomaly is present when one spinal region deviates from this typical number of vertebrae (Schmorl & Junghans, 1968).
Interestingly, segmentation anomalies are more likely to occur in the caudal part of the spine compared to the cranial spine. As such, the cervical count is highly conserved (Bronn, 1874; Le Double, 1912; Pilbeam & Young, 2004; Todd, 1922), with aberrations being often associated with detrimental effects, including late-term miscarriages and stillbirths (Galis et al., 2006; Schmorl & Junghans, 1968; ten Broek et al., 2012). Variation is more frequently observed at the thoracolumbar border and particularly the lumbosacral border, although the number of presacral vertebrae remains remarkably stable, both within humans and within mammals (Haeusler et al., 2002; Pilbeam, 2004; Schultz & Straus, 1945; Todd, 1922; Williams & Russo, 2015). In general, meristic changes, that is, additional or missing segments (Baur, 1891; von Ihering, 1878; Williams et al., 2019; Zuckerman, 1938), are less common than homeotic changes, which manifest as the transformation of one segment into a different segment (Barnes, 1994; Keith, 1902; Rosenberg, 1875, 1899; ten Broek et al., 2012; Williams et al., 2019). Such homeotic transformations shift the boundaries between spinal regions. The vertebra that assumes characteristics of two adjacent spinal regions is often referred to as a transitional vertebra (or “Zwischenwirbel,” Dürr, 1860).
Hox genes offer a genetic explanation for these homeotic changes since they encode for proteins controlling axial patterning and play a major role in the specification of the morphological identity of the vertebrae (Carapuço et al., 2005; Mallo et al., 2010; Müller et al., 2010). Thus, Hox11 is primarily responsible for the correct genesis of the sacral and caudal vertebrae. A complete lack of Hox11 gene products results in sacral vertebrae that are morphologically identical to lumbar vertebrae. Hox10 gene products are also important because of their rib-suppressing activity. Simultaneous inactivation of Hox11 and Hox10 leads to vertebrae bearing ribs in the lumbar and sacral spine (Mallo et al., 2010). Overexpression of Hox11, on the other hand, leads to signs of sacralization by fusion of adjacent vertebrae and a cranial shift of the first sacral vertebra (Carapuço et al., 2005).
Homeotic border shifts have also been implicated in the evolution of the spinal segmentation formulae of mammals and particularly of hominins (e.g., Haeusler et al., 2002, 2011; Machnicki & Reno, 2020; McCollum et al., 2010; Pilbeam, 2004; Williams et al., 2016, 2019; Williams & Pilbeam, 2021). However, the precise mechanisms responsible for the number and morphological identities of the vertebrae are still elusive (Kudlicki, 2019; Tague, 2018), and it is unknown whether these Hox genes are responsible for segmentation anomalies since Hox gene mutations also involve severe perturbation of limb and pelvic morphology (Wellik & Capecchi, 2003).
At the lumbosacral junction, border shifts can result in the last lumbar vertebra partly or completely fusing to the sacrum, resulting in a condition termed sacralization. In contrast, a partial or complete detachment of the first sacral element is referred to as lumbarization, in which case the corresponding vertebra morphologically resembles a last lumbar vertebra. The presence of transitional lumbar vertebrae has been linked to an increased risk for degenerative spine disorders, disc herniation, and low back pain (e.g., Bertolotti, 1917; Bron et al., 2007; Castellvi et al., 1984; Matson et al., 2020; Nardo et al., 2012; Vergauwen et al., 1997). Moreover, the patient's neurological symptoms may be at odds with the findings of Magnetic Resonance Imaging (MRI) examination, and spine surgery may consequently be performed at the incorrect segment if metameric variation is not considered.
Given their clinical relevance, segmentation anomalies have been extensively studied using X-ray, CT, and MRI imaging modalities (e.g., Hahn et al., 1992; Hughes & Saifuddin, 2006; Tins & Balain, 2016). For instance, vascular structures such as the right renal and the superior mesenteric arteries, the aortic bifurcation, the inferior vena cava confluence, and the celiac trunk have been investigated in terms of their spatial relationship to specific vertebrae in order to evaluate the presence and identity of transitional vertebrae (Lee et al., 2007; Ralston et al., 1992). Further, the origin of the iliolumbar ligament, which usually attaches at the costal process of the fifth lumbar vertebra (L5) (Hughes & Saifuddin, 2004, 2006; Lee et al., 2007), as well as vertebra and disc shape (Hsieh et al., 2000; O'Driscoll et al., 1996) have been also used for classifying transitional vertebrae. Nevertheless, most authors agree that analyzing the entire spine and counting the vertebrae from the atlas (Hahn et al., 1992; Hughes & Saifuddin, 2004; Tins & Balain, 2016), or at least from T12 (Bron et al., 2007 and references therein), is necessary to establish a reliable diagnosis, while successfully distinguishing sacralizations from lumbarizations remains challenging.
Despite their clinical importance, few studies offer reliable classification systems for the different stages of sacralizations and lumbarizations (Castellvi et al., 1984; Mahato, 2013; Tini et al., 1977). In fact, most studies (see Table 1) focus exclusively on sacralization while ignoring lumbarization, or they fail to distinguish between the two, thereby propagating further confusion in the literature. Additionally, some studies do not clearly state how the distinction was made or fail to consider the complete spinal count. This accounts for the broad range reported for the prevalence of segmentation anomalies, which varies between 4% and 35% (see Table 1, adapted from Bron et al., 2007). Even less attention has been dedicated to segmentation anomalies at the sacrococcygeal border. In fact, sacralization, that is, the synostosis of the first coccygeal element to the last sacral vertebrae, is a frequent occurrence and also results in a change in sacral element number (Lee et al., 2016; Tague, 2011; Woon et al., 2013).
TABLE 1. Previously published frequencies of transitional vertebrae (TV) from clinical studies and anatomical collections (studies based on archaeological material were omitted but see Drew & Kjellström, for an overview) Publication Data Sample origin n Lumbosacral transitional vertebrae Sacralizations Lumbarizations n % n % n % Andrew (1954) X-ray Great Britain 300 31 10.3 Apazidis et al. (2011) X-ray United States 211 75 35.5 Benlidayi et al. (2015) X-ray Turkey 1588 96 6.0 Cadeddu et al. (1997) X-ray United States 299 16 5.4 Castellvi et al. (1984) X-ray United States 200 60 30.0 Chaijaroonkhanarak et al. (2006) Osteological material Thailand 206 9 4.4 9 4.4 Chithriki et al. (2002) MRI United States 441 37 8.4 22 5.0 15 3.4 Dar and Peled (2014) CT Israel 436 57 13.1 Delport et al. (2006) X-ray United States 300 90 30.0 Elster (1973) CT United States 2000 140 7.0 Erken et al. (2002) X-ray Turkey 729 262 35.9 French et al. (2014) X-ray Australia 5429 540 9.9 225 4.1 315 5.8 Gopalan and Yerramshetty (2018) X-ray India 596 145 24.3 125 21.0 20 3.4 Hahn et al. (1992) MRI United States 200 24 12.0 15 7.5 9 4.5 Hald et al. (1995) X-ray Germany (males only) 10,922 792 7.3 850 7.8 650 6.0 Hsieh et al. (2000) X-ray United States 1668 67 4.0 Hughes and Saifuddin (2006) MRI Great Britain 500 67 13.4 46 9.2 21 4.2 Kim (1997) MRI Korea 690 41 5.9 12 1.7 29 4.2 Leboeuf et al. (1989) X-ray Australia 530 61 11.5 29 5.5 32 6.0 Lee et al. (2007) MRI South Korea 534 127 23.8 74 13.9 53 9.9 Luoma et al. (2004) MRI Finland (males only) 163 43 26.4 Mahato (2010) Osteological material India 330 20 6.1 Nardo et al. (2012) X-ray United States 4636 841 18.1 O'Driscoll et al. (1996) MRI Great Britain 100 15 15.0 Otani et al. (2001) MRI/CT Asia 1009 119 11.8 Peh et al. (1999) MRI China 129 17 13.2 8 6.2 9 7.0 Peterson et al. (2005) X-ray Canada 353 43 12.2 Quinlan et al. (2006) MRI Ireland 769 35 4.6 Santiago et al. (2001) CT Spain 138 26 18.8 16 11.6 10 7.2 Steinberg et al. (2003) X-ray Israel 464 85 18.3 65 14.0 20 4.3 Tague (2009) Osteological material United States 2086 131 6.3 Tang et al. (2014) X-ray China 5860 928 15.8 0.0 Taskaynatan et al. (2005) X-ray Turkey 881 48 5.4 40 4.5 8 0.9 Tini et al. (1977) X-ray Switzerland 4000 269 6.7 Tins and Balain (2016) MRI Great Britain 418 14 3.3 8 1.9 4 1.0 Uçar et al. (2013) X-ray Turkey 3607 683 18.9 17.2 1.7 Vergauwen et al. (1997) CT Belgium 350 53 15.1 Total 53,192 6146 12The correct recognition of the exact type of segmentation anomaly is crucial to understanding its relationship to disc herniation, degenerative joint disorders, and low back pain. Moreover, lumbarization cannot reliably be distinguished from sacralization in fragmentary skeletal remains, thereby obscuring any potential functional implications caused by such anomalies. Understanding the morphological variation, that these anomalies introduce, has important implications in both clinical and archaeological contexts.
In this study, we evaluate morphological characteristics associated with segmentation anomalies of the modern human sacrum. Specifically, we aim to establish repeatable methods that reliably distinguish sacralizations from lumbarizations, while emphasizing informative morphological changes at the base of the sacrum, namely the diagnostic relationship between the iliac crest and the linea terminalis that co-occur with segmentation anomalies.
2 MATERIALS AND METHODSOur modern human sample consisted of 145 clinical CT scans, of which 127 included the entire vertebral column (n = 51) or a complete thoracolumbosacral spine (n = 76). Resolution ranged from 0.5 to 2.0 mm, with slice thicknesses being 0.8–3.0 mm. The sample comprised 88 patients aged 14.5–20.0 years from the Hôpital de la Timone, Marseille, and 58 adults aged 22–47 from the Department of Biomedical Imaging and Image-guided Therapy, Medical University of Vienna, Vienna. The individuals were analyzed in two age cohorts (subadults and adults) based on the extent of closure at the primary ossification centers (about 20 years). The patients from both the Viennese and French datasets were randomly selected from larger data sets, provided they did not suffer any developmental or skeletal growth disorders. The French data were anonymized according to the personal privacy standards of the French National Ethical Committee (Corron, 2016). The Viennese data were anonymized by the Data Clearing House of the Medical University of Vienna, Austria, after clearance by the ethic commission of the Medical University of Vienna (votum 1196/2017).
The datasets were segmented using Amira (www.fei.com), and surface models were generated to represent the sacrum, the hip bones, the lumbar and the thoracic vertebrae, as well as the costae. The cranialmost rib-bearing vertebra was classified as the first thoracic vertebra (T1). Vertebrae at the thoracolumbar border with rudimentary ribs were recorded as either thoracic (if the rib was mobile and oriented caudally) or as lumbar (if the rib was broad and blunt and shaped like a detached costal process directed laterally or cranially; see Haeusler et al., 2002). A laterally oriented costal process warranted a lumbar classification. Vertebrae with asymmetrical transitions were counted as half belonging to one spinal region and half belonging to the other spinal region. Thus, sacral vertebrae were counted according to the number of sacral foramina divided by two plus one (Schultz & Straus, 1945). Lumbosacral transitional vertebrae were classified according to Castellvi et al. (1984).
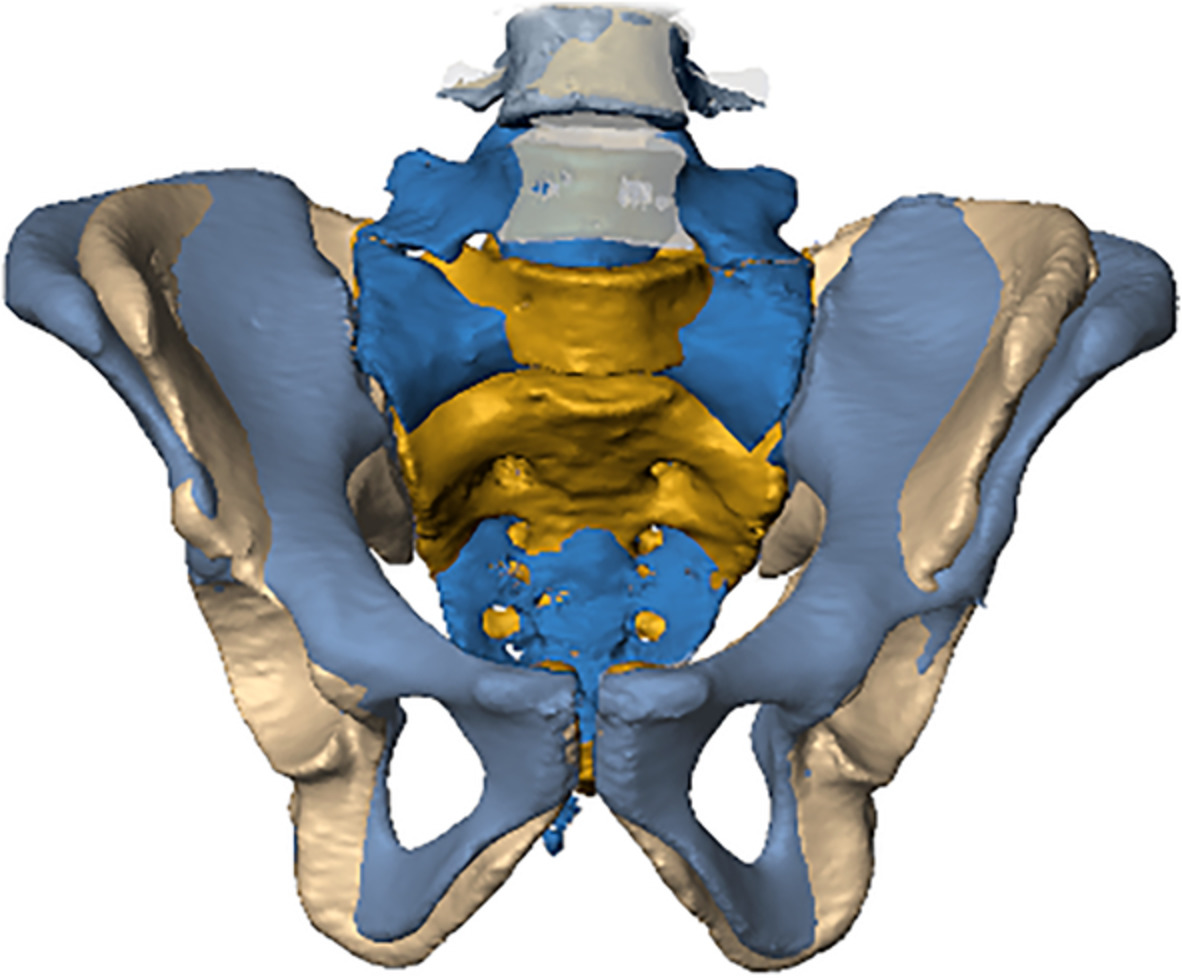
Additionally, we used the iliac crest (Andrew, 1954; Farshad-Amacker et al., 2015) and the linea terminalis (Tague, 2009) as references to identify the type of segmentation anomaly (i.e., sacralization or lumbarization, Figure 1). For that purpose, we established four different categories for the relationship between the iliac crest and L4/L5, with the level of the iliac crest ranging between (1) the center of L4, (2) the inferior surface of L4, (3) the superior surface of L5, and (4) the center of L5. Levels were recorded from the 3D models generated from the clinical CTs in the supine position. Similarly, three different configurations were recognized for the trajectory of the linea terminalis. Specifically, the trajectory of the margin of the pelvic brim onto the sacral alae (the iliac portion of linea terminalis) could either be (1) upwards, (2) straight, or (3) downwards with respect to the arcuate line (Figure 1). Moreover, we recorded the location of the promontory, that is, that part of the spinal column that was most anteriorly protruding into the pelvic inlet, either sacral or lumbar, and the presence of a second promontory (Lierse, 1987). Associations between all variables, including sex, were investigated using χ2 and Cramérs V using SPSS (www.ibm.com).

(a) Drawing of the lower spine and the pelvis in anterior view illustrating the spatial relationships between the iliac crest and the fourth and fifth lumbar vertebrae, the linea terminalis, which is composed of the arcuate line and the margin of the pelvic brim on the ala of the first sacral vertebra (marked as “ala margin”). (b) Visualization of the categorial relationship between the iliac crest and the fourth (L4) and fifth (L5) lumbar vertebrae, namely (1) center of L4, (2) inferior surface of L4, (3) superior surface of L5, and (4) center of L5. (c) The trajectories from the arcuate line on the hipbones to the margin of the pelvic brim of the sacral alae. The pelvic inlet thereby forms a flat circular plane (2) but can also be elongated, bent upwards (1), or curtailed and bent downwards (3)
We also recorded aberrations at the sacrococcygeal level. The sacrococcygeal articulation is a cartilaginous joint reinforced by numerous sacrococcygeal ligaments. At the sacrococcygeal border, sacralizations of the coccyx or coccygealizations, that is, detachment of the fifth sacral vertebra, have rarely been investigated (Lee et al., 2016; Tague, 2011; Woon et al., 2013; Woon & Stringer, 2012). Nevertheless, sacralizations of the first coccygeal vertebra are quite common, either
留言 (0)