
記住我

Neurodegenerative disease and metabolic disorders share many pathophysiological features, including disruption of cellular energy metabolism and inflammation. As such, midlife occurrence of cardiometabolic disorders such as type II diabetes mellitus, obesity, dyslipidaemia, hypertension, atherosclerosis and cardiovascular disease significantly increase risk for future diagnosis of Alzheimer's disease (AD) at ages 65+.1, 2 However, these risk factors are modifiable—and therapeutic management of them is shown to reduce the increased risk they confer.3 Therefore, uncovering the mechanistic links between cardiometabolic disorders and neurodegenerative disease may illuminate new, desperately needed strategies to combat the escalating incidence of AD.4
While cardiometabolic diseases typically originate in peripheral tissues/organ systems, their chronic effects eventually affect the central nervous system.5, 6 Neurons are among the most metabolically demanding cells of the body and require extensive energy resources for communication and survival; therefore, deficits in cellular energy metabolism profoundly affect brain function and structure.7 Compounding these problems, chronic peripheral inflammation associated with cardiometabolic disorders contributes to inflammatory changes in the brain, promoting protein misfolding and accumulation of extracellular amyloid beta (Aβ)8 and intracellular neurofibrillary tau tangles.9 In turn, these hallmark proteinopathies of AD trigger immune-mediated responses from microglia and astrocytes, exacerbating neuroinflammation and neurodegeneration.10-12
Irisin, a hormone released by myocytes and other cells (including neurons), may be capable of therapeutically addressing metabolic dysfunction, inflammation and neuropathology.13 Discovered in 2012, the glycoprotein irisin is a cleaved fragment of the transmembrane fibronectin type III domain-containing protein 5 (FNDC5).13 When the skeletal muscle is subject to exercise, FNDC5 is cleaved into a soluble irisin fragment, which is then secreted into the bloodstream.13-15 FNDC5 and irisin are also constitutively expressed in other oxidative tissues/organs such as bone, liver, heart and brain.13, 16-19 Peripherally, irisin is shown to restore energy homeostasis and optimise cellular energy utilisation by ‘browning’ white adipose tissue and enhancing thermogenesis13, resolving insulin resistance and improving glucose utilisation in diabetic models20, 21, and reducing inflammation22 and expression of inflammatory genes.23 Irisin is thought to mediate the beneficial effects of exercise on the brain. Exercise-induced FNDC5/irisin release enhances expression of brain-derived neurotrophic factor (BDNF) in the hippocampus, promoting synaptic plasticity, hippocampal neurogenesis and memory formation.24-26 Compelling data show that irisin may provide neuroprotection against ischaemic damage after stroke27 and neurodegenerative diseases such as multiple sclerosis28 and AD.29 Irisin treatment has been used to rescue memory deficits, restore synaptic impairments, and ameliorate Aβ-induced suppression of BDNF in APP/PS1dE9 AD-model mice30; reduce soluble Aβ oligomer formation in cultured APP/PS1 neurons30, 31; prevent cleavage of amyloid-precursor protein (APP) into aggregation-prone Aβ42 fragments31; and ameliorate neuroinflammation.32 Notably, no studies have yet investigated how irisin affects the hyperphosphorylation of tau (ptau) and its subsequent assembly into damaging intraneuronal neurofibrillary tangles (collectively known as tauopathy), another major neuropathological hallmark of AD and other forms of dementia. The goal of this study was to determine whether pre-treatment with exogenous irisin protein could reduce the emergence of early ptau accumulation and inflammation in the brains of transgenic htau mice, a selective model of age-related tauopathy.
MATERIALS AND METHODS SubjectsMale and female htau mice (B6.Cg-Mapt tm1(GFP) Tg(MAPT)8cPdav/J; Jax stock# 005491) on a C57BL/6 J background were used for the primary analyses of this study. Htau mice are a tau overexpression model that was created by random insertion of the human microtubule-associated protein tau (MAPT) gene into ‘tau knockout’ mice that have a neomycin cassette-induced disruption at exon 1 of the endogenous mouse Mapt.33, 34 The human MAPT transgene is driven by a tau promoter that causes these mice to overexpress all six isoforms (including 3R/4R) of human MAPT in the absence of endogenous mouse Mapt.33, 34 Phenotypically, htau mice form age-related accumulation of hyperphosphorylated tau (ptau) that leads to microtubule destabilisation, ptau aggregation and formation of neurofibrillary tangles in the hippocampus and frontal cortices, among other brain areas.33, 34 This tauopathy coincides with significant neuroinflammation and contributes to age-related cognitive deficits in htau mice.33, 35 Htau mice have been characterised for studying dementia-related tauopathy analogous to late-onset AD and frontotemporal dementia.36 However, two caveats when considering the external validity of data obtained from this model are that htau mice produce tau isoforms in different ratios than humans,37 and the genomic integration site and conformation of the human MAPT/vector transgene array in this model have not yet been determined. Age- and sex-matched C57BL/6 J (C57) mice, the background strain of htau mice, were used as non-transgenic healthy controls in this study. For secondary analyses, an additional group of age- and sex-matched ‘tau null’ mice (littermates of htau) were included in the study. While these mice function as genetic and negative controls for their htau littermates, tau null mice lack both the humanised MAPT transgene and endogenous Mapt gene and are thus tau deficient—exhibiting phenotypic abnormalities across neural, metabolic and behavioural parameters 38, 39 that make them inadequate as primary healthy controls.
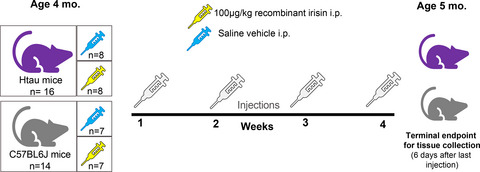
In total, 16 htau (8 per sex) and 14 C57 (7 per sex) mice were used for the primary analyses of this study. Data from female and male mice were not combined for analysis but evaluated in separate data sets 40 because of anticipated sex differences in htau phenotype based on our previous work,38, 41 as well as an increasing number of studies describing inherent sex differences in other AD models.42-44 Study treatment was initiated in mice at the ‘pre-symptomatic’ age of 4 months, where early isoforms of ptau and indicators of neuroinflammation are minimally present in in non-cognitive brain regions38, 41 and are just emerging in the hippocampus and frontal cortex.35 This pre-symptomatic staging was used to approximate the clinical relevance of mitigating early metabolic risk factors associated with AD as a preventative measure rather than a restorative treatment. Mice were 5 months old at the terminal endpoint of the study when tissue was collected for histology. A separate replicative cohort of age- and sex-matched tau null (n = 15) and htau (n = 13) mice were used to optimise irisin delivery and provide control data for relevant assays (see Figure S1).
All mice were bred at the Northeast Ohio Medical University (NEOMED) from original stocks obtained at Jax Mice; all groups were housed under identical conditions. All experimental procedures were approved by the NEOMED Institutional Animal Care and Use Committee and conducted in accordance with the Guide for Care and Use of Laboratory Animals published by the National Institutes of Health.
TreatmentFemale and male mice were stratified into treatment groups by body weight to ensure overall uniformity of subjects. Htau (n = 8/sex) and C57 (n = 7/sex) mice in the experimental treatment group received one weekly intraperitoneal (i.p.) injection of 100 µg/kg recombinant human irisin protein (r-irisin; #11451: Cayman Chemicals; Ann Arbor, MI) reconstituted in sterile saline at a concentration of 10 µg/ml. Dosage, frequency and route of administration for r-irisin were based on previous studies that demonstrated reproducible treatment effects.18, 45 Htau (n = 8/sex) and C57 (n = 7/sex) mice in the vehicle control group received i.p. injections of sterile saline. Prior to injection, mice were weighed and individually coded syringes were pre-loaded with a weight-based volume of r-irisin or vehicle to keep experimenters blind to treatment during the actual injection procedure. Mice received a total of 4 weekly injections across one month's time, with injections being delivered at the same time on the same day each week. A timeline for treatment administration, experimental groups and ages is presented in Figure 1. Group numbers and experimental parameters for the secondary control replicative cohort of tau null and htau mice are provided in Figure S1.

Illustration of timeline and experimental groups in study. Four-month-old 'pre-symptomatic’ female and male htau and matched non-AD control (C57BL/6 J) mice were assigned to irisin and vehicle treatment groups. Mice received weekly injections of either 100 µg/kg recombinant human irisin or equivalent volume of saline vehicle for the duration of 4 weeks. Six days after the last treatment injection, mice (at 5 months of age) were sacrificed for fresh tissue collection. Thirty mice were used for the primary analyses of this study with the strain/treatment group breakdown as follows: 16 htau mice: 8 per treatment group; 14 C57BL/6 J mice: 7 per treatment group. All groups had equivalent numbers of each sex so that female and male data could be analysed separately
Fresh tissue collection and analysis for proteinSix days after the last treatment injection, mice were sacrificed via decapitation under 4% inhaled isoflurane administered via anaesthetic vaporiser. The hippocampus, prefrontal cortices, brainstem and hypothalamus were freshly dissected, immediately frozen on dry ice and then stored at −80°C until further analysis. Trunk blood was collected and centrifuged for 10 min at 4°C (at 2000 g) for separation/extraction of serum which was then stored at −80°C until use. The freshly dissected hippocampus, brainstem, hypothalamus and prefrontal cortex were prepared for automated capillary-based western blotting on a Wes platform (Protein Simple). The hippocampus was targeted as the primary region of interest, because it is a major area impacted by neuropathology in htau mice and is also shown to express FNDC5.26 Secondary regions assessed included: brainstem, in which 4-month-old htau mice exhibited elevated ptau (serine 202) in serotonergic neurons of the dorsal raphe nucleus38; hypothalamus, a structure involved in metabolic regulatory functions that has been shown to express FNDC5/irisin45; and prefrontal cortex, a common target of tauopathy in older htau mice.35
Tissue preparation for Wes occurred as described previously,38 but briefly, brain tissue was homogenised in ice-cold T-PER buffer containing protease and (where relevant) phosphatase inhibitors (Thermo Scientific #78440) via sonification (10% amplitude for three 2-s pulses), and centrifuged at 14,000 rpm for 20 min at 4°C. The Wes platform detects and provides quantitative size-based measurement of targeted proteins by computing spectra of chemiluminescence signals versus the apparent molecular weight of protein, which are determined by mapping ladder peaks to capillary positions based on fluorescent signalling of labelled protein standards of known molecular weight.46 Wes was used to quantify tau (ptau ser202; Cell Signaling #11834; 1:75 and total tau; Tau5 Abcam#ab80579; 1:25), FNDC5 (ProteinTech #23995-1-AP; 1:25) and TNFα (Rockland Scientific #210-401-321S; 1:10) in brain homogenates. For quantification of ptau levels, we measured tau phosphorylated at serine 202 because this form is shown to emerge in the pre-symptomatic pathological stage we were targeting in htau mice,38 as well as in the incipient pathology stage in human clinical samples.35, 38, 47, 48 We chose TNFα as our surrogate marker of inflammation in both brain and peripheral circulation because it is a pro-inflammatory cytokine commonly observed in htau mice and shown to be mitigated by irisin.49 All antibodies were diluted (ProteinSimple antibody diluent 2; #042-195) prior to use. Tissue homogenates were loaded into 12–230 kDa (ptau, tau, FNDC5) and 2–40 kDa (TNFα) Wes plates for automated assay. To permit comparison with tau data from our previously published studies,38 GAPDH (Sigma #G9545; 1:5000) was used as the loading control for ptau and total tau assays; for other analytes (FNDC5 and TNFα), a total protein detection module (ProteinSimple, #DM-TP01) was used to control for the amount of protein loaded into the assay. Tissue homogenates from tau null mice were used as negative control samples for ptau plates. Recombinant irisin protein was loaded as a positive control for FNDC5/irisin plates. Compass software (ProteinSimple, v3.1.7) was used to perform peak area calculations using the default Gaussian method and generate artificial lane view images from the spectra for graphic representation. Compass provided molecular weight, signal intensity (area), percent area, and signal-to-noise readout from each sample in units of normalised chemiluminescence. ‘Virtual’ bands were generated by Compass software using chemiluminescence values to graphically represent signal molecular weight and intensity for each analyte and are provided in Figure S2.
Enzyme-Linked immunosorbent assay for measurement of irisin in serumIrisin was measured in serum samples from mice using a validated enzyme-linked immunosorbent assay (ELISA) kit (Phoenix Pharmaceuticals #EK-067–29: Burlingame) shown to measure the 12 kDA form of cleaved irisin.28, 50, 51 Serum was diluted 10-fold in assay buffer and both samples and positive controls were ran in duplicate per manufacturer instructions. Assay sensitivity was 1.84 ng/ml with a dynamic range of 0.1–1000 mg/ml. Intra- and inter-assay coefficients of variation were <10% and <15%, respectively. Assay absorbance was read at 450 nm on a Molecular Devices SpectraMax M5 plate reader and reported in ng/ml.
Statistical analysisGroup sizes were calculated with a priori power analysis using G*Power software52 based on our previously published effect sizes in htau mice.38, 41 Statistical analysis was performed using IBM SPSS 26 Software. Separate datasets were created for female and male mice so that each sex could be analysed independently of each other. This was done to increase the sensitivity of detecting treatment effects within each sex strata,40, 53 as the design of the study was not intended to directly compare the sexes by including sex as an independent variable. Prior to analysis, data were screened for outliers and normalcy using boxplots and frequency distributions in SPSS. Outliers that extended more than 2.5 standard deviations from subgroup mean were evaluated and in two cases where measurement error was obvious, data were removed. Factorial analysis of variance (ANOVA) models were built for each dataset (female and male) and were used to test for treatment (irisin; vehicle) x strain (htau; control) effects and interactions. To follow-up significant interactions in each data set, data were stratified (using the SPLIT function in SPSS) by treatment to test for strain differences and then by strain to test for treatment effects within these strata. Bonferroni-corrected post hoc ANOVA were used to determine subgroup differences. In the absence of significant interactions, simple main effects were used to describe group differences. Pearson correlations were used to describe relationships between continuous variables to provide additional information. Data from the replicative cohort with tau null and htau mice were analysed separately; statistics and results are provided in legend of Figure S1.
RESULTSStatistical results for omnibus factorial ANOVAs are provided in Table 1.
TABLE 1. Main effect and interaction statistics for omnibus ANOVAs conducted on sex-stratified datasets Factorial ANOVA F, p and eta2 values Effect Ptau HC TNFα HC TNFα serum FNDC5 HC Irisin serum Females (n = 15) df 1, 7 1, 7 1, 10 1, 7 1, 11 Strain 59.44, p < 0.001* 5.187, p = 0.042* 74.54, p < 0.001* 3.332, p = ns 0.005, p = ns Treatment 9.03, p = 0.02*, eta2 = 0.099 8.273, p = 0.014*, eta2 = 0.233 11.06, p < 0.001*, eta2 = 0.051 0.262, p = ns 21.16, p = 0.001*, eta2 = 0.656 Interaction 11.30, p = 0.012*, eta2 = 0.124 6.458, p = 0.026*, eta2 = 0.182 86.36, p < 0.001*, eta2 = 0.40 0.165, p = ns 0.059, p = ns Males (n = 15) df 1, 7 1, 7 1, 10 1, 7 1, 11 Strain 21.87, p = 0.002* 106.89, p < 0.001* 22.66, p < 0.001* 14.44, p = 0.007* 0.072, p = ns Treatment 0.061, p = ns 9.016, p = 0.017*, eta2 = 0.066 0.255, p = ns 1.517, p = ns 34.11, p < 0.000*, eta2=0.747 Interaction 0.056, p = ns 13.585, p = 0.006*, eta2 = 0.099 0.187, p = ns 0.091, p = ns 0.077, p = ns Strain = htau or C57 control; treatment = r-irisin or vehicle control; interaction = strain by treatment comparison. For sex, F = female; M = male; HC = hippocampus. FNDC5 HC analyses reported above based on composite variable created from sum of 20 and 37kDa forms. Degrees of freedom (df), ANOVA statistic (F), significance value (p) and effect sizes (eta2) are provided for treatment main effects and interactions. Hippocampal ptau levels are significantly reduced in female htau mice after 4 weeks of exogenous irisin treatmentFigure 2 summarises protein assay data for ptau and total tau measured from brain samples of htau mice via automated western blotting. Female htau mice treated with r-irisin exhibited a significant reduction in hippocampal ptau (F1,8 = 9.82, p = 0.01; Figure 2A). This finding represented a 30% difference in ptau levels between vehicle-treated and r-irisin treated htau mice after only 4 weeks of injections. Total tau was not affected by treatment (Figure 2B). Ptau did not significantly differ as a function of treatment group in brainstem, hypothalamus, or prefrontal cortex (Figure 2C).

Mean tau (ptau serine 202 and total tau) chemiluminescence in brain regions of htau mice as a function of irisin treatment. (A) Female htau mice treated with r-irisin (striped bars) had a significant reduction in hippocampal ptau202 compared to vehicle-treated (solid bars) conspecifics. (B) Irisin treatment did not have an effect on total tau levels in female htau hippocampus. (C) Ptau in female htau prefrontal cortex, brainstem and hypothalamus did not statistically differ as a function of treatment. (D) In male htau mice, r-irisin treatment had no effect on ptau. (E) Irisin treatment did not have an effect on total tau levels in male htau hippocampus. (F) Ptau level did not vary by treatment in any other brain regions of male htau mice. Both ptau202 and total tau data were calibrated for the amount protein loaded per sample using peak intensity chemiluminescence data for GAPDH. Error bars show s.e.m.; * indicates significant treatment group difference. p < 0.05 in all significant comparisons. n = 16 htau mice (4/sex/treatment group)
Recombinant irisin treatment does not reduce neural ptau load in male htau miceIrisin treatment had no effect on ptau levels in male htau mice in any brain region (Figure 2D,F), nor did it affect total tau (Figure 2E).
Irisin treatment significantly reduces neuroinflammation in female htau miceIrisin-treated female htau mice had significantly reduced hippocampal TNFα relative to their vehicle-treated counterparts (F1,5 = 12.97, p < 0.05; Figure 3A). Notably, TNFα and ptau levels in the hippocampus were significantly correlated in female htau mice (Figure 3B). While not statistically significant, TNFα levels in female htau brainstem and prefrontal cortex showed a trend towards similar irisin-related reductions (Figure 3B). Hippocampal TNFα levels were significantly greater in untreated female htau mice compared to C57 controls (Figure 3A) and tau null controls (Figure S1), but this difference was eliminated in irisin-treated htau mice.

TNFα as a surrogate marker of inflammation in brain and serum of mice. Mean TNFα chemiluminescence in brain and serum of htau (purple) and control C57BL/6 J (grey) mice as a function of treatment (striped bars = r-irisin treated mice; solid bars = vehicle-treated mice). (A) Treatment with r-irisin significantly reduced elevated levels of pro-inflammatory cytokine TNFα in hippocampus of female htau mice. TNFα in C57 control mice did not vary as a function of treatment. (B) Brainstem and prefrontal cortex ptau levels appeared to be reduced in irisin-treated female htau mice, but this was not statistically significant. Scatterplots depict strong correlation (indicated by Pearson's r value, p < 0.05) between hippocampal TNFα and ptau measurements for individual female htau mice. (C) Mean serum measurements of TNFα were significantly reduced in irisin-treated female htau mice. (D) Irisin-treated male htau mice showed significantly enhanced hippocampal inflammation compared to vehicle-treated htau mice. (E) TNFα also appeared to be elevated in irisin-treated male htau prefrontal cortex data but this was not statistically significant. Scatterplots illustrate the lack of correlation between hippocampal TNFα and ptau measurements for individual male htau mice (p = ns). (F) Mean serum measurements of TNFα did not differ as a function of irisin treatment in male htau mice, however they trended towards an irisin-induced increase. Results are calibrated by total protein loaded per sample. Error bars show s.e.m.; * indicates significant difference between treatment groups. # indicates significant strain difference between htau and C57 mice of same treatment group. p < 0.05 in all significant comparisons. n = 16 htau mice (4/sex/treatment group); 14 C57 mice (3-4/sex/treatment group)
Irisin treatment exacerbated neuroinflammation in male htau miceIrisin treatment produced different results in male htau mice; it enhanced hippocampal TNFα levels compared to male htau mice given saline vehicle, (F1,5 = 30.63, p = 0.003; Figure 3D). Unlike female htau mice, male htau hippocampal TNFα levels were not correlated with overall neural ptau load (Figure 3E). TNFα levels in the prefrontal cortex of r-irisin-treated male htau mice also appeared to be slightly elevated relative to vehicle-treated mice (Figure 3E), but this was not statistically supported. Hippocampal TNFα levels were significantly greater in male htau mice compared to C57 controls (Figure 3D) and tau null controls (Figure S1) regardless of treatment condition.
Irisin treatment effects on htau peripheral inflammation were consistent with effects on htau neuroinflammationSince r-irisin was administered peripherally via intraperitoneal injection, we measured serum levels of TNFα to determine whether this treatment affected systemic inflammation. Consistent with our findings in neural tissue, r-irisin treatment reduced serum TNFα levels in female htau mice, (F1,6 = 16.25, p = 0.007; Figure 3C), but failed to do so in male htau mice (Figure 3F). While not meeting criteria for statistical significance, it was noted that serum TNFα levels showed a trend of being higher among r-irisin treated males compared to those that received the saline vehicle, (F1,4 = 5.8, p = 0.07, ns).
FNDC5/irisin levels in the brain did not change as a function of irisin treatmentNeural levels of FNDC5/irisin were measured to determine whether peripherally administered irisin directly increased these protein levels in the brain. We detected several bands of FNDC5/irisin at different molecular weights (Figure S2)—a finding consistent with previously published literature that identified multiple FNDC5/irisin bands associated with the significant amount of post-translational modification/glycosylation of this protein that occurs in the brain.17, 30, 54, 55 Therefore, we quantified and reported data from two bands (20 kDA and 37 kDA) that were consistent with those reported by Varela-Rodríguez et al. 45 that represented un-glycosylated and glycosylated forms of FNDC5, respectively. Post-translational modification also causes the molecular weight range to overlap between the full-length FNDC5 membrane-bound protein and truncated irisin.54, 56 Both are shown to colocalise in parenchymal brain tissue, preventing specific determination of the cleaved protein (irisin) from its parent (FNDC5) in our Wes assays. Therefore, to preserve accuracy, we refer to these measurements in brain tissue as ‘FNDC5/irisin’ as was done by Lourenco and colleagues.30 In contrast, our ELISA measurements in serum are presumed to contain only the cleaved, secreted irisin protein and not membrane-bound FNDC5, thus we refer to these as ‘irisin’.30, 57
For initial comparisons, we used a composite-dependent variable for FNDC5/irisin created from the sum of both 20 and 37 kDa values for each brain region. The results did not indicate any significant treatment-related changes in composite FNDC5/irisin (Figure 4A,B), but in male mice, there was a significant strain effect for the hippocampus (Table 1), prefrontal cortex (F1,8 = 8.48, p = 0.02) and brainstem (F1,8 = 18.28, p = 0.004). To further evaluate these findings, we analysed the 20 and 37 kDa bands separately. Data from vehicle-control mice showed that male htau had intrinsically elevated neural 37kDA FNDC5/irisin compared to male C57 mice in the hippocampus (F1,3 = 83.79, p = 0.003; Figure 4B), prefrontal cortex (F1,4 = 11.65, p = 0.03), hypothalamus (F1,4 = 12.24, p = 0.03) and brainstem (F1,3 = 23.31, p = 0.02)—a region which also showed significantly increased measurements of the 20 kDA form (F1,3 = 36.88, p = 0.009). Data are summarised in Figure 4C.

Irisin levels in mice. Mean FNDC5/irisin levels (37kDA band) in the hippocampus of htau (purple) and control C57BL/6 J (grey) mice as a function of treatment (striped bars = r-irisin treated mice; solid bars = vehicle-treated mice). (A) Female brain FNDC5/irisin levels did not differ as a function of treatment or strain. (B) Male brain FNDC5/irisin levels did not differ as a function of treatment, but vehicle-treated male htau mice exhibited significantly higher FNDC5/irisin levels than vehicle-treated C57 males. (C) Dot density scatterplots show individual composite measurements summed across 20kDA and 37kDA bands for FNDC5/irisin protein in the hippocampus, brainstem, hypothalamus and prefrontal cortex for female and male htau and C57 mice. These summed composite values were normalised across all brain regions for depiction. (D&E) Serum levels of irisin are elevated in female and male mice treated with r-irisin regardless of strain. * indicates significant treatment group differences, significance = p < 0.05. n = 16 htau mice (4/sex/treatment group); 14 C57 mice (3–4/sex/treatment group)
Serum irisin levels were elevated in response to treatment conditionContrary to measurements in brain tissue, serum irisin levels were significantly enhanced in both female and male mice receiving r-irisin treatment regardless of strain (F1,22 = 51.39, p < 0.01; Figure 4D,E), confirming that peripheral administration of 100 µg/kg r-irisin effectively doubled the amount of circulating irisin in mice. This finding also showed that blood levels of irisin were sustained for a relatively long duration post-injection, as serum samples were collected from mice six days after the last treatment. No intrinsic differences in circulating irisin levels were shown between mouse strains nor did these levels appear to differ between females and males.
DISCUSSIONPrevious studies support a protective role for irisin against AD-associated amyloid pathology and inflammation.30, 31 Here, we expand this knowledge on irisin's therapeutic potential with evidence that one month of peripherally delivered irisin significantly reduced hippocampal ptau and neuroinflammation in female htau mice—a model that approximates (but is not identical) to tauopathy in clinical dementia. This reduction was observed in pre-symptomatic female mice at an age where they only exhibit ‘pre-tangle’ and emerging pathology—therefore, it remains to be determined how irisin impacts more advanced aspects of tauopathy including neurofibrillary tangle formation and human pathology. However, these findings provide important insights to irisin's potential as an early intervention in the disease process.
It is notable that the therapeutic effects of irisin were specific to female htau mice. Our data showed that irisin treatment did not alter ptau levels in male htau brains and actually seemed to enhance inflammation in these mice. While we anticipated some degree of sex differences in our results, this large divergence in findings was unexpected. AD disproportionately affects women4 by reasons that are not fully understood; a lack of direct, sex-based characterisations of both AD clinical presentations and experimental model phenotypes has likely contributed to this.58 While an increasing number of studies include subjects of both sexes, sex is not always analysed as a variable and data are often pooled for analysis. This is problematic for the field; if we had pooled data across sexes in this study, we would have obscured the intriguingly dichotomous sex-based treatment effects of irisin (recall Figures 2 & 3).
As such, clinical and experimental studies that analyse sex-stratified data or directly test for sex effects are beginning to identify large sex differences in pathological phenotype.41-44, 59 Sex differences in irisin levels have been shown in human serum and cerebrospinal fluid samples, although the functional significance of this remains unknown.51, 60, 61 FNDC5 gene polymorphisms have been linked to cardiovascular complications in diabetic women but not men,62 suggesting that alterations in basal levels of FNDC5/irisin may confer sex-based vulnerability to metabolic dysfunction. Sex differences in circulating gonadal hormones may also influence response to irisin. For example, gonadectomy was shown to alter ser
留言 (0)