
記住我



The seasonally dry tropical forests (SDTFs) of Latin America make an important contribution to Neotropical plant species diversity. High levels of species endemism across numerous geographically separate SDTF areas (Pennington et al., 2009) result in high total species numbers (DRYFLOR, 2016). However, this diversity is currently highly threatened because in most areas less than 10% of the original SDTF extent remains (García et al., 2014). Some SDTF nuclei are large (Caatinga in northeastern Brazil; Misiones in Paraguay and northern Argentina; and Chiquitano in eastern Bolivia; Dirzo et al., 2011), but smaller areas occur in inter-Andean pockets from Bolivia to Colombia and Venezuela (DRYFLOR, 2016; Särkinen et al., 2012).
SDTFs in inter-Andean valleys contribute to the outstandingly high plant species richness that characterizes the Andean biodiversity hotspot (DRYFLOR, 2016). Our understanding of the origin and evolutionary assembly of Andean SDTFs primarily derives from the central Andes (i.e., Bolivia, Peru, and southern Ecuador). A series of phylogenetic studies of plant clades found in dry valleys in the Central Andes revealed patterns of strong phylogenetic geographic structure (e.g., Killeen et al., 2007; Luebert & Weigend, 2014; Pennington et al., 2010; Särkinen et al., 2012). These patterns are most clearly evidenced in phylogenies that sample multiple individuals per species, which have revealed monophyletic species and monophyletic populations within species that are confined to individual dry valleys. Within these valley-confined species and populations, sequence divergence is often low, but the lineages themselves are ancient—often 5–10 million years old (e.g., Pennington et al., 2010). Such long stem lineages and shallow crown groups are consistent with the persistence of small populations over evolutionary timescales in these valleys. This persistence also suggests long-term isolation of these individual dry valley SDTFs, with little successful lineage immigration, making them analogous to islands (Flantua et al., 2020; Särkinen et al., 2012).
Andean orogeny and erosion formed the SDTF valleys. The habitat within those valleys must have assembled when the surrounding mountain range (“cordillera”) was sufficiently high to create the topographic effects that cause the characteristic seasonal and overall low annual rainfall (ca. 1,800 mm and a 5–6 month long dry season; Dexter et al., 2015; Killeen et al., 2007; Arias et al., 2021). Understanding Andean orogeny is therefore central to inferring the assembly of the Andean forest biomes. In the central Andes, different phases of uplift occurred >45 million years ago (Ma) and at 17–10 Ma, while the modern central Andes topography was formed over the past 5.4 Ma (Garzione et al., 2017). The northern Andes comprise three different cordilleras. Topographic growth of the central cordillera started during the Cretaceous, with major phases of uplift during the Miocene (Zapata et al., 2020), and in the western cordillera, it started in the Eocene and increased during the middle Miocene (León et al., 2018). The orogeny of the eastern cordillera started ca. 30 Ma (Bayona et al., 2021; Cooper et al., 1995; Horton et al., 2020; Mantilla Figueroa et al., 2016), which created positive relief since at least 20 Ma (Boschman, 2021; Horton et al., 2010; Ochoa et al., 2012; Siravo et al., 2018), and reached modern elevation at least by 3 Ma (Mora et al., 2020; Torres et al., 2005; Van der Hammen et al., 1973). This geological history predicts a later assembly of dry forest in the smaller and scattered SDTF pockets in the northern Andes (younger forests in inter-Andean pockets of northern Ecuador, Colombia, and Venezuela), and therefore, we predict the stem age of lineages endemic there to be less than 3 Ma.
Here, we evaluate whether the age of a species endemic to northern Andean SDTFs is consistent with the timing of the Andean orogeny in this region. We do this by inferring the evolutionary history of Cavanillesia chicamochae Bombacoideae: Malvaceae (Fernández Alonso, 2003), an emblematic succulent-trunked tree found only in the Chicamocha Canyon of Colombia. Cavanillesia is a genus of four species, found in different biomes in Central and South America.
The Bombacoideae have repeatedly evolved specialization to SDTFs (Zizka et al., 2020), and in Cavanillesia, C. chicamochae is the only species endemic to SDTF. While C. umbellata is found in the SDTF of northeastern Brazil, it also grows in the rain forests of western Amazonia. Cavanillesia is characterized by nocturnal flowers that are generally pollinated by hawk moths and bees, and whose seeds that are dispersed by wind (Murawski et al., 1990). Cavanillesia chicamochae is specifically pollinated by Apoideae (Hymenoptera), Lycenidae and Nymphalidae (Lepidoptera), and hummingbirds (Trochilidae; Díaz-Pérez et al., 2011). There is a documented importance of the mucilage in seed germination and drought resistance in Cavanillesia (Garwood, 1985; Wolfe, 2017), and the seasonal rains cause the seeds of C. chicamochae to become covered with up to 5 cm of mucilage, which serves as a water reserve and ensures successful germination in the dry Chicamocha Canyon (Díaz-Pérez et al., 2011; Fernández Alonso, 2003).
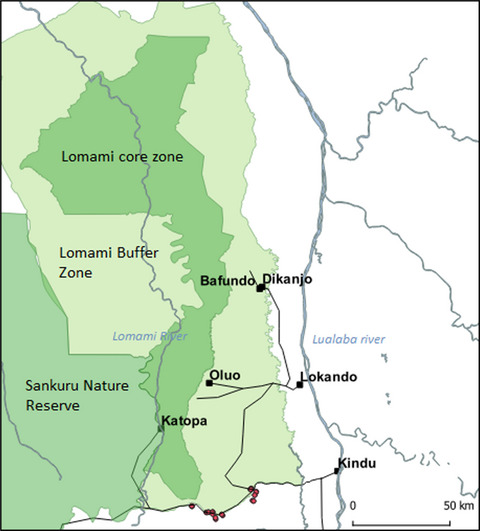
The Chicamocha, located in the eastern cordillera of the Andes, is the second largest canyon on Earth (Figure 1b). However, the SDTF of the Chicamocha Canyon, where C. chicamochae is endemic, has been little explored and is under severe threat due to livestock grazing and deforestation (Díaz-Pérez et al., 2011; Pizano & García, 2014). In particular, goats are identified as the root cause of disturbance in the area, and in response C. chicamochae is characterized as endangered (Lopez-Gallego & Morales, 2020). At least six plant species are endemic to the SDTFs of the Chicamocha Canyon (Albesiano & Fernández Alonso, 2006; Albesiano & Rangel-Ch, 2006; Albesiano et al., 2003; Díaz-Pérez et al., 2011; Fernández Alonso, 2014).
 Geographic distribution of Cavanillesia samples used in this study from northern South America. (a) Symbols denote different species, as specified in the legend. Filled symbols represent samples used in the phylogenetic analyses, and open symbols represent occurrence data derived from the Global Biodiversity Information Facility. The georeferenced localities for each species reflect their approximate present-day geographic range. The outlined green areas show the current distribution of seasonally dry tropical forests (SDTF; DRYFLOR, 2016). Geologically, the central and northern Andes are divided by the Amotape-Huancabamba depression formed by the Marañón River in northern Peru (Horton, 2018), shown here with a black bracket. (b) The Chicamocha Canyon and distribution of sampling localities, with open triangles replaced with crosses to enhance visualization of geographically dense samples. Blue arrows indicate the direction of river water flow, where all rivers shown flow into the Sogamoso, which drains into the Magdalena River and then into the Caribbean Sea
Geographic distribution of Cavanillesia samples used in this study from northern South America. (a) Symbols denote different species, as specified in the legend. Filled symbols represent samples used in the phylogenetic analyses, and open symbols represent occurrence data derived from the Global Biodiversity Information Facility. The georeferenced localities for each species reflect their approximate present-day geographic range. The outlined green areas show the current distribution of seasonally dry tropical forests (SDTF; DRYFLOR, 2016). Geologically, the central and northern Andes are divided by the Amotape-Huancabamba depression formed by the Marañón River in northern Peru (Horton, 2018), shown here with a black bracket. (b) The Chicamocha Canyon and distribution of sampling localities, with open triangles replaced with crosses to enhance visualization of geographically dense samples. Blue arrows indicate the direction of river water flow, where all rivers shown flow into the Sogamoso, which drains into the Magdalena River and then into the Caribbean Sea
Our understanding of the historical assembly of Andean SDTF forests is limited by a lack of data on the biogeographic patterns in the northern Andes. We present results based on sampling multiple individuals per species from multiple populations in the dry inter-Andean valleys of the northern Andes. By also sampling multiple populations of Cavanillesia chicamochae across the Chicamocha Canyon and all its congeneric species, we ask: (i) is the age of C. chicamochae consistent with the timing of the northern Andean orogeny, implying that mountain-building created the seasonally dry forest habitat in the Chicamocha canyon?; and (ii) is there genetic differentiation amongst populations within the Chicamocha Canyon? Answering these questions will both fill a knowledge gap in our understanding of the biogeographic history of Andean SDTFs and provide information of relevance to the conservation of these highly threatened ecosystems.
2 METHODSPhylogenetic relationships within Cavanillesia were assessed using dense taxon sampling of all species and 39 accessions of C. chicamochae. We conducted fieldwork throughout the Chicamocha Canyon area of Santander, Colombia, and three of the larger populations of Cavanillesia chicamochae were sampled for this work (Figure 1). We also collected material of other Cavanillesia species from Bolivia, Brazil, Colombia, Costa Rica, and Peru (Figure 2).

Photographs of three of the four known Cavanillesia species. (a) C. chicamochae (Photo: Christine D. Bacon); (b) C. platanifolia (Photo: Jeferson Sobrinho Cavalho); (c) C. umbellata (Photo: Jeferson Sobrinho Cavalho)
DNA was extracted from silica-gel dried leaf tissue using DNeasy plant kits (QIAgen). DNA sequences for ITS (Baum et al., 1998), trnL-trnF (Taberlet et al., 1991), and trnK-matK (Nyffeler et al., 2005) and ETS (Andreasen & Baldwin, 2001) were sequenced at Macrogen (Korea). All new sequences generated for this study have been deposited in GenBank under accession numbers KM245161- KM245261 (Appendix S1).
As outgroups, sequences of the above loci for all genera in the core Bombacoideae were downloaded from GenBank, as well as representatives of the sister group, Malvatheca, following the taxonomy of Duarte et al. (2011).
Nucleotide alignments were obtained independently for each of the four loci using default parameters using the Muscle (Edgar, 2004) tool in Geneious v7.0.5 (Biomatters Ltd.) and manual adjustments were performed following Simmons (2004). Data for 77 individuals and 964 bp of nuclear and plastid data were partitioned by locus to allow for variation in substitution models, which can alleviate or remove the impact of saturation caused by using relatively older fossils for inferences directed at more recently evolved nodes (sensu Ho et al., 2005).
Divergence times were estimated by constraining the phylogeny at the crown node of the Malvatheca clade, which corresponds to the stem node of Bombacoideae (Duarte et al., 2011), using Malvaciphyllum macondicus fossil leaves (Carvalho et al., 2011) from the Cerrejón Paleocene forests of Colombia (Wing et al., 2009). Carvalho et al. (2011) used synapomorphies and unique character combinations to support natural affinities for the fossil leaves, which allow for robust placement in the phylogeny. We used an exponential prior for the age of the calibrated node, defined by a mean of 0.7 and an offset value of 58.0 to account for uncertainty in the geological age of the fossil deposit and to infer temporal evolution.
The dated phylogenetic analysis was run using BEAST v1.8.0 (Drummond et al., 2006; Drummond & Rambaut, 2007) with an uncorrelated lognormal molecular clock model, a Yule pure birth speciation model with no starting tree, the GTR +Γ model of nucleotide substitution with four rate categories, and the default operator. Markov chains were run for 100 million generations and repeated three times to test for convergence and to ensure effective sample sizes exceeded 200. After verifying that chains reached stationarity in Tracer v1.6, 25% was removed as burn-in and files were combined in LogCombiner v1.8.0. Tree files were combined to estimate the median node height and the 95% highest posterior density was estimated in TreeAnnotator v1.8.0.
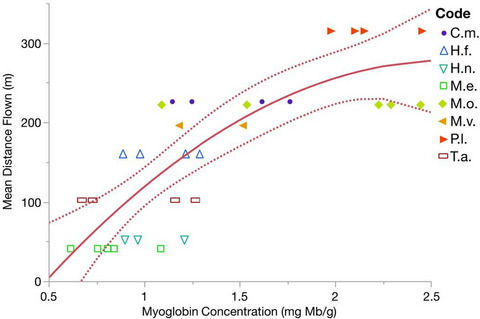
3 RESULTS AND DISCUSSIONThe crown age of Cavanillesia is estimated at a mean of 8.8 Ma (Figure 3), which is consistent with the age of several other SDTF-endemic plant genera or clades from the central Andes and Neotropical SDTFs more generally (Pennington et al., 2004, 2010; Särkinen et al., 2012). All Cavanillesia species and each of the populations of C. chicamochae were resolved as monophyletic and recovered with high posterior probability in our Bayesian analysis (Figure 4). Our phylogenetic reconstruction suggests two main patterns: (1) species relationships are not predicted by geographic proximity, and (2) relationships may be better predicted by ecological preference because the sister species C. chicamochae and C. umbellata are the only two species found in SDTF. These results fit prior patterns of niche conservatism found in Neotropical SDTF (e.g., Särkinen et al., 2012).

Phylogeny of the Bombacoideae with mean ages of divergence times, geological time scale, and the position of the fossil used to calibrate the tree (Photo: Monica Carvalho)
 Phylogeny of Cavanillesia showing posterior probabilities along branches, where symbol shapes and colors match those in Figure 1
Phylogeny of Cavanillesia showing posterior probabilities along branches, where symbol shapes and colors match those in Figure 1
The mean stem age of C. chicamochae is 1.46 Ma (highest posterior density age range 2.06–0.45 Ma; Table 1), which is within the margin of uncertainty of the most recent uplift phase of the eastern cordillera of Colombia (5 to 2 Ma; Cediel et al., 2003; Gregory-Wodzicki, 2000; Horton et al., 2020; Boschman, 2021), but may be too young to explain diversification as a function of Andean orogeny itself. C. chicamochae is younger than species from the western cordillera; legumes from the Valle de Cauca region between the western and central cordilleras have a SDTF-endemic species that originated ca. 4 Ma (Leutzelburgia jacana; Trabuco da Cruz et al., 2018). The age of Cavanillesia chicamochae is consistent with the Chicamocha endemic Niceforo's Wren (Thryophilus nicefori), which diverged c. 1.7 Ma (Lara et al., 2012). Together, these results suggest a later timing of colonization of the SDTF of the Chicamocha Canyon area by plants and animals than that of central Andean valleys such as the Marañon, Peru, where endemic plant genera, or clades within genera, have stem ages between 27 and 5 Ma (Särkinen et al., 2012). The younger ages found in the Chicamocha canyon are consistent with a later origin of the SDTF there.
TABLE 1. Credible intervals of absolute age estimation each of the Cavanillesia species, with age in millions of year (Ma) Species Stem age Crown age C. chicamochae 1.46 (2.06–0.45) 1.11 (1.41–0.40) C. hylogeiton 7.24 (11.11–2.76) 0.87 (1.25–0.00) C. platanifolia 3.73 (5.24–1.80) 0.55 (0.70–0.07) C. umbellata 1.46 (2.06–0.45) 0.72 (1.00–0.02)The three populations of C. chicamochae sampled were each resolved as monophyletic with 1.0 PP, showing strong population structure within the species (Figure 4). This phylogeographic pattern is consistent with that shown for the central Andes, where populations of the legume Cyathostegia mathewsii were resolved as monophyletic. However, in that case, populations were sampled in deep valleys separated by high cordillera (Loja, Ecuador; Marañon and Apurimac/Mantaro, Peru) and coalescence (i.e., monophyly) had been achieved in time periods of 5.4 – 2.8 Ma (Pennington et al., 2010). In contrast, the populations of C. chicamochae separated over the last 1 Ma. Populations of different bird species also showed strong population structure, developed as recently as over the last 0.25 Ma (Arbeláez-Cortés & Trujillo-Arias, 2021). The coalescence of individual populations within the restricted Chicamocha area suggests considerable dispersal limitation within the SDTF of the canyon. This result underscores that the limitation of immigration and establishment by a lack of resident plant mortality in these drought-adapted dry forests may be as important in shaping their biogeography as any physical barriers such as mountain cordilleras or oceans (Pennington & Lavin, 2015). Our results support the hypothesis that genetic coalescence (i.e., population and or species monophyly) within short time periods is achieved in areas where population sizes are small and dispersal and subsequent population establishment is limited (i.e., dry Andean valleys; Pennington & Lavin, 2015).
Multiple changes to the landscape and their influence in driving local climatic and vegetational heterogeneity likely interacted to create and maintain strong population structure in C. chicamochae over relatively small spatial scales. This likely happened as a result of physical and ecological barriers preventing gene flow between populations. Some of these changes occurred before the diversification of populations, while others happened simultaneously with it. The eastern cordillera experienced mountain uplift and exhumation over the last 20 Ma, which increased since the Plio-Pleistocene (Boschman, 2021; Siravo, Faccenna, et al., 2019; Siravo et al., 2018). A recent geomorphological study revealed that the fault system related to the Chicamocha Canyon has caused tectonic activity and intense land sliding and changing river drainage patterns (García-Delgado et al., 2019). Further erosion leading to the extant configuration of the canyon dates to the Pleistocene (Caballero, Mora, et al., 2013; Caballero, Parra, et al., 2013; Julivert, 1958). These landscape changes during the Pleistocene likely contributed to the genetic subdivision of C. chicamochae.
The precipitation regime over the Chicamocha region is driven by a complex interaction between the annual movement of the Intertropical Convergence Zone (ITCZ), atmospheric moisture advection by low-level jets from the Choco, Caribbean, and Orinoco regions, and land surface-atmospheric interactions (Arias et al., 2021; Hoyos et al., 2018). The origination of C. chicamochae occurred during the Middle Pleistocene Transition, after which colder, longer and drier glacial periods set in (Clark et al., 2006) that could have increased the aridity over the Chicamocha region, contributing to the formation of SDTF. Taken together, the formation of the canyon and its local climatic conditions created pockets of SDTF that waxed and waned through time in response, causing strong population structure within C. chicamochae. Our results do not indicate whether SDTF was once widespread across the Chicamocha Canyon region in the past or not, but certainly today the distribution of C. chicamochae is patchy (Figure 1b).
Our results for C. chicamochae have important conservation implications. This species, together with other Chicamocha Canyon endemics, indicate that effective management of this region is required to conserve its unique biodiversity (Collazos-González et al., 2020). Over nine million hectares of SDTF were once found in Colombia, but after deforestation and desertification across the country, only 8% remain and only 5% are included within the Colombian National System of Protected Areas (SINAP; Pizano & García, 2014). In 2020, efforts in Colombia were made to protect SDTFs in nearby regions (e.g., Distrito Regional de Manejo Integrado Bosque Seco Tropical Pozo Azul, Norte de Santander department), but the Chicamocha Canyon is not currently included in SINAP. Species inventories (e.g., Albesiano & Fernández Alonso, 2006; Albesiano & Rangel-Ch, 2006; Albesiano et al., 2003; Fernández Alonso, 2014) are a critical first step for identifying areas of high species diversity and high endemism (Kattan et al., 2019). However, design and management of protected areas is fundamental and should involve collaboration among scientists, policymakers, politicians, and—perhaps most importantly—local communities. Protecting areas in the Chicamocha Canyon would help meet the Aichi biodiversity target 11 of the Convention on Biological Diversity, calling for conservation of 17% of terrestrial areas of importance for biodiversity, but critically, our results showing clear genetic differentiation of C. chicamoachae within the Chicamocha Canyon suggest that multiple reserves are needed to protect the genetic diversity of this species.
4 CONCLUSIONSHere, we show a middle Quaternary of origin of Cavanillesia chicamochae (1.46 Ma), a SDTF species endemic to the Chicamocha Canyon, that could be explained by either a very young orogeny of the eastern Andes or climatic changes in the Middle Pleistocene Transition. This age is younger than that of plant species endemic to the central Andes, as predicted by the later orogeny of the area. Our new data from the poorly studied northern Andean SDTFs suggest a general pattern of plant biogeography of SDTFs across the Andes that emphasizes dispersal limitation leading to species and even population monophyly within individual dry valleys. These results, together with the presence of other endemic plant and bird species, clearly advocate for conservation areas in the Chicamocha Canyon area. Critically, the clear genetic differentiation of populations of C. chicamochae within the small area of the Chicamocha Canyon implies that multiple protected areas will be necessary to conserve the full genetic diversity of this species.
ACKNOWLEDGMENTSWe thank J. Hernández Torres, K. Banda-R., S. Faurby, H. Hooghiemstra, M. Landis, F. Luebert, A. Mora, A. Rojas, G. Romero, T. Särkinen, E. Tesón, E. Tenorio, and F. Velásquez-Puentes for support and discussions; J.G. Carvalho-Sobrinho, B. Jestrow, K. Dexter, D. Díaz, and C. Maldonado for providing material; and the editor and anonymous reviewers whose feedback greatly improved the quality of this work. Open access funding enabled and organized by Projekt DEAL
CONFLICT OF INTERESTThe authors declare no conflict of interest related to this work.
The data (DNA sequences) that support this study are openly available at NCBI (https://www.ncbi.nlm.nih.gov/genbank/) with the reference numbers found in Appendix 1.
Filename Description btp13031-sup-0001-AppendixS1.pdfPDF document, 84.3 KB Appendix S1Please note: The publisher is not responsible for the content or functionality of any supporting information supplied by the authors. Any queries (other than missing content) should be directed to the corresponding author for the article.
REFERENCES
Albesiano, S., & Fernández Alonso, J. L. (2006). Catalogue of the vascular plants from the Chicamocha river canyon (tropical zone), Boyacá-Santander, Colombia. First Part. Caldasia, 28(1), 23– 44. Albesiano, S., & Rangel-Ch, J. O. (2006). Structure of the plant communities from Chicamocha canyon, 500–1200 masl; Santander, Colombia: a tool for conservation. Caldasia, 28(2), 307– 325. Albesiano, S., Rangel-Ch, J. O., & Cadena, A. (2003). Vegetation of the Chicamocha River Canyon (Santander, Colombia). Caldasia, 25(1), 73– 99. Andreasen, K., & Baldwin, B. G. (2001). Unequal evolutionary rates between annual and perrenial lineages of checker mallows (Sidalcea, Malvaceae): evidence from 18S–26S rDNA internal and external transcribed spacers. Molecular Biology and Evolution, 18(6), 936– 944. Arbeláez-Cortés, E., & Trujillo-Arias, N. (2021). Role of the Chicamocha River Canyon on the phylogeography of humid montane forest birds in Colombia. Journal of Avian Biology, e02833. https://doi.org/10.1111/jav.02833 Arias, P. A., Garreaud, R., Poveda, G., Espinoza, J. C., Molina-Carpio, J., Masiokas, M., Viale, M., Scaff, L., & van Oevelen, P. J. (2021). Hydroclimate of the Andes part II: Hydroclimate variability and sub-continental patterns. Frontiers in Earth Science. https://doi.org/10.3389/feart.2020.505467 Baum, D. A., Small, R. L., & Wendel, J. F. (1998). Biogeography and floral evolution of baobabs (Adansonia, Bombacaceae) as inferred from multiple data sets. Systematic Biology, 47(2), 181– 207. https://doi.org/10.1080/106351598260879 Bayona, G., Baquero, M., Ramírez, C., Tabares, M., Salazar, A. F., Nova, G., Duarte, E., Pardo, A., Plata, A., Jaramillo, C., Rodríguez, G., Caballero, V., Cardona, A., Montes, C., Gómez-Marulanda, S., & Cárdenas-Rozo, A. (2021). Unraveling the widening of the earliest Andean northern orogen: Maastrichtian to early Eocene intra-basinal deformation in the northern Eastern Cordillera of Colombia. Basin Research, 33(1), 809– 845. Boschman, L. M. (2021). Andean mountain building since the Late Cretaceous: A paleoelevation reconstruction. Earth-Science Reviews, 220, e103640. https://doi.org/10.1016/j.earscirev.2021.103640 Caballero, V., Mora, A., Quintero, I., Blanco, V., Parra, M., Rojas, L. E., Lopez, C., Sánchez, N., Horton, B. K., Stockli, D., & Duddy, I. (2013). Tectonic controls on sedimentation in an intermontane hinterland basin adjacent to inversion structures: The Nuevo Mundo syncline, Middle Magdalena Valley, Colombia. Geological Society, London, Special Publications, 377(1), 315– 342. https://doi.org/10.1144/SP377.12 Caballero, V., Parra, M., Mora, A., López, C., Rojas, L. E., & Quintero, I. (2013). Factors controlling selective abandonment and reactivation in thick-skin orogens: A case study in the Magdalena Valley, Colombia. Geological Society, London, Special Publications, 377(1), 343– 367. https://doi.org/10.1144/SP377.4 Carvalho, M. R., Herrera, F. A., Jaramillo, C. A., Wing, S. L., & Callejas, R. (2011). Paleocene Malvaceae from northern South America and their biogeographical implications. American Journal of Botany, 98(8), 1337– 1355. https://doi.org/10.3732/ajb.1000539 Cediel, F., Shaw, R. P., Cáceres, C., & Bartolini, C. (2003). Tectonic assembly of the Northern Andean block. In C. Bartolini, R. T. Buffler, & J. Blickwede (Eds.), The Circum-Gulf of Mexico and the Caribbean: Hydrocarbon Habitats, Basin Formation and Plate Tectonics (Vol. 79, pp. 815– 848). Tulsa, USA: American Association of Petroleum Geologists. Clark, P. U., Archer, D., Pollard, D., Blum, J. D., Rial, J. A., Brovkin, V., Mix, A. C., Pisias, N. G., & Roy, M. (2006). The middle Pleistocene transition: Characteristics, mechanisms, and implications for long-term changes in atmospheric CO2. Quaternary Science Reviews, 25(23), 3150– 3184. Collazos-González, S. A., Zuluaga-Carrero, J., & Cortés-Herrera, J. O. (2020). Birds of the Chicamocha Canyon: a call for conservation. Biota Colombiana, 21(1), 58– 85. Cooper, M. A., Addison, F. T., & Álvarez, R. (1995). Basin development and tectonic history of the Llanos Basin, Eastern Cordillera and Middle Magdelena Valley, Colombia. AAPG Bulletin, 79(10), 1421– 1443. Dexter, K. G., Smart, B., Baldauf, C., Baker, T. R., Balinga, M. P. B., Brienen, R., Fauset, S., Feldpausch, T. R., Silva, L.-D., Muledi, J. I., Lewis, S. L., Lopez-Gonzalez, G., Marimon-Junior, B. H., Marimon, B. S., Meerts, P., Page, N., Parthasarathy, N., Phillips, O. L., Sunderland, T., … Pennington, R. T. (2015). Floristics and biogeography of vegetation in seasonally dry tropical regions. International Forestry Review, 17(S2), 10– 32. https://doi.org/10.1505/146554815815834859 Díaz-Pérez, C. N., Puerto-Hurtado, M. A., & Fernández-Alonso, J. L. (2011). Evaluación del hábitat, las poblaciones y el estatus de conservación del barrigón (Cavanillesia chicamochae, Malvaceae-Bombacoideae). Caldasia, 33(1), 105– 119. Dirzo, R., Young, H. S., Mooney, H. A., & Ceballos, G. (2011). Seasonally dry tropical forests: Ecology and conservation. Springer. Drummond, A. J., Ho, S. Y. W., Phillips, M. J., & Rambaut, A. (2006). Relaxed phylogenetics and dating with confidence. PLoS Biology, 4(5), 699– 710. https://doi.org/10.1371/journal.pbio.0040088 Drummond, A. J., & Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology, 7, e214. https://doi.org/10.1186/1471-2148-7-214 DRYFLOR (2016). Plant diversity patterns in neotropical dry forests and their conservation implications. Science, 353(6306), 1383– 1387. https://doi.org/10.1126/science.aaf5080 Duarte, M. C., Esteves, G. L., Salatino, M. L. F., Walsh, K. C., & Baum, D. A. (2011). Phylogenetic analysis of Eriotheca and related genera (Bombacoideae, Malvaceae). Systematic Botany, 36(3), 690– 701. Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32(5), 1792– 1797. https://doi.org/10.1093/nar/gkh340 Fernández Alonso, J. L. (2003). Bombacaceae neotropicae novae vel minus cognitae VI. Novedades en los géneros Cavanillesia, Eriotheca, Matisia y Pachira. Revista De La Academia Colombiana De Ciencias Exactas, 27(102), 25– 37. Fernández Alonso, J. L. (2014). Salvia guaneorum (Labiatae), a new species from the Chicamocha Canyon, Colombia. Phytotaxa, 156, 221– 228. https://doi.org/10.11646/phytotaxa.156.4.4 Flantua, S. G. A., Payne, D., Borregaard, M. K., Beierkuhnlein, C., Steinbauer, M. J., Dullinger, S., Essl, F., Irl, S. D. H., Kienle, D., Kreft, H., Lenzner, B., Norder, S. J., Rijsdijk, K. F., Rumpf, S. B., Weigelt, P., Weigelt, P., & Field, R. (2020). Snapshot isolation and isolation history challenge the analogy between mountains and islands used to understand endemism. Global Ecology and Biogeography, 29(10), 1651– 1673. García, H., Corzo, G., Isaccs, P., & Etter, A. (2014). In C. Pizano, & H. García (Eds.), DISTRIBUCIÓN Y ESTADO ACTUAL DE LOS REMANENTES DEL BIOMA DE BOSQUE SECO TROPICAL EN COLOMBIA: INSUMOS PARA SU GESTIÓN. El Bosque Seco en Colombia (pp. 228– 251). Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. García-Delgado, H., Villamizar-Escalante, N., & Bernet, M. (2019). Recent tectonic activity along the Bucaramanga Fault System (Chicamocha River Canyon, Eastern Cordillera of Colombia): a geomorphological approach. Zeitschrift Für Geomorphologie, 62(3), 199– 215.
留言 (0)