
記住我

The incidence and mortality of lung cancer rank first in the world, and more than 80% of patients are nonsmall cell lung cancer (NSCLC).1 More than 50% of NSCLC patients are in the middle or advanced stage when they are first diagnosed, and they have lost the opportunity for surgical treatment. The traditional treatments usually focus on radiotherapy and chemotherapy, and the results are often unsatisfactory. The appearance of small molecule tyrosine kinase inhibitors (TKIs) that target EGFR, such as gefitinib, is a major advance in NSCLC treatment in recent years, and they become the first line treatment of NSCLC patients with sensitive mutations of EGFR. However, these patients relapse after 9–13 months of continuous use of the drug, that is, secondary drug resistance.2 Although the application of the irreversible TKI inhibitors afatinib and AZD9291, which specifically target the EGFR T790M mutation, can partially alleviate resistance, it can still produce new site resistance mutations. Therefore, research on the mechanism of EGFR TKI resistance is in the ascendant, and there is a long way to go to find further targets related to EGFR TKIs resistance.
Long noncoding RNA (long noncoding RNA) is a type of noncoding RNA with a length greater than 200 nt, and its relationship with tumor chemotherapy resistance has become a research hotspot in recent years. Many studies have confirmed that lncRNA is closely related to drug sensitivity. For example, lncARSR could up-regulate AXL and c-MET by competitively binding miR-34/miR-449 to induce sunitinib resistance in renal cancer cells. At the same time, lncARSR also transmitted drug-resistant phenotypes through exosomes. The drug-resistant phenotype could be transmitted through exosomes which may become a potential therapeutic target for sunitinib resistance.3 The miR-100 and miR-125b produced by lncRNA MIR100HG synergistically inhibit five Wnt/β-catenin negative regulators, activate the Wnt signaling pathway, and thereby mediate cetuximab resistance.4 These studies suggest that the interaction between noncoding RNAs may play an important role in tumor resistance. Therefore, further searching for new targets for gefitinib resistance from the perspective of lncRNA will help provide new strategies for the treatment of lung cancer resistance.
We continuously induced the gefitinib-sensitive cell line HCC827 and successfully established the secondary drug-resistant cell line HCC827/GR.5 In this study, we screened out the lncRNAs related to gefitinib resistance through microarray data. This study will help reveal the role and mechanism of lncRNA in the secondary resistance of gefitinib and provide a new therapeutic target for reversing the secondary resistance of EGFR-TKI.
2 MATERIALS AND METHODS 2.1 Reagents and cell cultureThe human NSCLC cell line HCC827 is sensitive to gefitinib and gefitinib-resistant cell line (HCC827/GR) was generated as described previously.5 The cells were cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Waltham, Massachusetts) with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific), 100 U/ml penicillin, and 50 μmol/L β-mercaptoethanol (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The cells were in an incubator containing 5% CO2 at 37°C.
2.2 Cell transfectionThe small interfering RNAs (siRNAs) that specifically target human lncRNA ZMIZ1-AS1 were purchased from RiboBio Co., Ltd. For transfection, the cells were placed in six-well or 96-well plates. Twenty-four hours later, they were transfected with lncRNA ZMIZ1-AS1 silencer or negative control (NC) by Lipofectamine® RNAiMAX reagent (cat. no. 13778075) and Gibco® Opti-MEM® (cat. no. 31985062) according to the manufacturer's instructions. The cells were treated with or without different concentrations of gefitinib for 48 h.
2.3 Cell viability assayThe CCK8 assay was used to detect cell viability. The cells were placed at approximately 5 × 103 cells per well in a 96-well plate and stimulated with different concentrations of gefitinib after 24 h of cell attachment. CCK8 reagents were added into the wells after 48 h and the OD values (absorbance) were measured at 450 nm using a Microplate Reader (BioTek ELx800; BioTek Instruments, Winooski, Vermont).
2.4 Cell apoptosis analysisThe cells were plated at 1 × 106 cells/well in a six-well plate followed by gefitinib treatment. Twenty-four hours later, the cells were stained using a fluorescein isothiocyanate/Annexin V apoptosis detection kit (Cat.no. 556547; BD Biosciences, San Diego, California). The samples were then loaded onto a flow cytometer (C6; Becton Dickinson, San Diego, California).
2.5 Quantitative real-time PCRTotal RNA was isolated using TRIzol reagent (Invitrogen). And then, the cDNA was synthesized from 100 ng extracted total RNA using the PrimeScript RT reagent Kit (Takara Bio Company, Shiga, Japan) and amplified by real-time (RT)-qPCR with The PowerUp™ SYBR® Green Master Mix (Takara Bio Company) on an ABI 7300 System (Applied Biosystems, Foster City, California). The PCR was performed under the following conditions: (a) 94°C for 30 s; (b) 40 cycles of 94°C for 5 s and 60°C for 30 s; and (c) 95°C for 1 min, 55°C for 30 s, and then 95°C for 30 s. The primers for the lncRNA were designed and purchased from RiboBio Co. Ltd. Each experiment was performed in triplicate and ACTB (encoding β-actin) was selected as a house-keeping gene. The relative gene expression levels were determined by 2−ΔΔCt method.
2.6 RNA pull-down assayA Magnetic RNA Protein Pull-Down Kit (Pierce) was used for RNA pull-down assays according to the manufacturer's instructions. Biotinylated ZMIZ1-AS1 RNA were synthesized by RiboBio (China). For each assay, 50 pmol biotinylated RNA were incubated with 50 μl prewashed with streptavidin Dynabeads (Invitrogen) for 1 h at 4°C. RNA-bound beads were then incubated with lysates from HCC827/GR cells cytosolic/nuclear extracts and eluted proteins were further separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and the gel was silver stained. Then, lncRNA ZMIZ1-AS1 specific bands were subjected to mass spectrometry and retrieved in human proteomic library. hnRNP A1 antibody (4B10; sc-32301) was used in western blot analysis.
2.7 Statistical analysisAll the figures were prepared using GraphPad Prism 8.0.2 software (GraphPad Inc., La Jolla, California). Spss software (version 20.0; IBM Corp., Armonk, New York) was performed for statistical analysis. The comparison of cell viability and apoptosis between multiple groups was performed using two-way analysis of variance (ANOVA). The differences between two groups were analyzed using student's t test. Differences with p < .05 were considered statistically significant.
3 RESULTS 3.1 LncRNA ZMIZ1-AS1 was increased in gefitinib resistance cellsCell viability was determined by the CCK-8 assay to assess the resistance index of the gefitinib resistance cells. As shown in Figure 1A, the IC50 value of HCC827 for gefitinib was 0.05 μM. However, the IC50 of HCC827/ GR became >10 μM. The resistance index was more than 200-fold higher. The microarray experiment in gefitinib-sensitive HCC827 and gefitinib-resistant HCC827/GR cells in pairs was performed as described previously.6 The microarray data have been submitted to the Gene Expression Omnibus (accession number, GSE74575). We found that ZMIZ1-AS1 was significantly upregulated in HCC827/GR cells by screening the microarray data. Further RT-qPCR results were consistent with the microarray data (Figure 1B).

The expression of ZMIZ1-AS1. (A) Validation of gefitinib-resistant cell line. (B) Real-time quantitative PCR results of lncRNA ZMIZ1-AS1. **p < .01
3.2 ZMIZ1-AS1 knockdown significantly attenuated gefitinib resistanceTo elucidate the role of ZMIZ1-AS1 in gefitinib sensitivity of HCC827/GR cells, HCC827/GR cells were transfected with ZMIZ1-AS1 siRNA or NC siRNA. Following 48 h treatment, ZMIZ1-AS1 expression was measured using RT-qPCR. The results demonstrated that ZMIZ1-AS1 expression was significantly downregulated in cells transfected with ZMIZ1-AS1 siRNA compared with cells transfected with NC siRNA (Figure 2A). ZMIZ1-AS1 siRNA significantly decreased HCC827/GR cell viability following gefitinib stimulation compared with NC group (Figure 2B). Cells were analyzed using flow cytometry and the results of the apoptosis assay were similar to those of the CCK-8 assay. The apoptosis rates were 3.13 ± 0.21, 3.60 ± 0.27, 16.57 ± 0.83, and 31.33 ± 0.27 in the HCC827/GR + NC siRNA, HCC827/GR+ ZMIZ1-AS1 siRNA, HCC827/GR + NC siRNA +gefitinib, and HCC827/GR+ ZMIZ1-AS1 siRNA +gefitinib groups, respectively (Figure 2C). Transfection with ZMIZ1-AS1 siRNA prior to gefitinib exposure significantly increased the sensitivity of HCC827/GR cells to gefitinib compared with the cells pretreated with NC siRNA. These results indicate that ZMIZ1-AS1 can influence the sensitivity of HCC827/GR cells to gefitinib.
 The effect of ZMIZ1-AS1 on gefitinib resistance. (A) HCC827/GR cells were transiently transfected with NC siRNA or ZMIZ1-AS1 siRNA. Unpaired Student's t tests were used in Figure 2A. The transfected cells were then treated with gefitinib for a further 48 h. Treated cells were harvested for a cell viability assay (B) and flow cytometry (C). Statistical comparisons were performed using two way ANOVA. Data are presented as means ± SD.**p p
3.3 ZMIZ1-AS1 can bind to hnRNPA1
The effect of ZMIZ1-AS1 on gefitinib resistance. (A) HCC827/GR cells were transiently transfected with NC siRNA or ZMIZ1-AS1 siRNA. Unpaired Student's t tests were used in Figure 2A. The transfected cells were then treated with gefitinib for a further 48 h. Treated cells were harvested for a cell viability assay (B) and flow cytometry (C). Statistical comparisons were performed using two way ANOVA. Data are presented as means ± SD.**p p
3.3 ZMIZ1-AS1 can bind to hnRNPA1
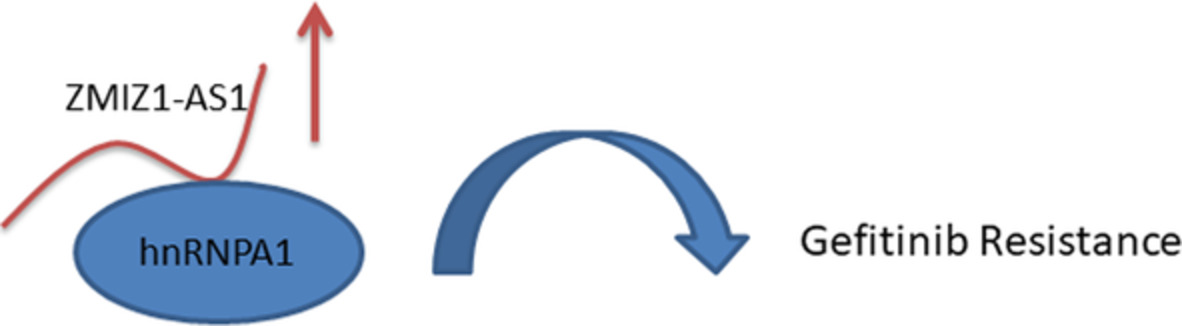
To explore the potential binding proteins of ZMIZ1-AS1in HCC827/GR cells, we performed a biotin-labeled RNA pull-down assay followed by silver staining (Figure 3A). A protein band specifically presented in lncRNA-HGBC was located at approximately >35 kD and then was subjected to sequence analysis via mass spectrometry. With great interest, we paid particular attention to an RNA-binding candidate hnRNPA1 that has high confidence score. Western blot assay confirmed that hnRNPA1 was a specific binding protein for ZMIZ1-AS1 (Figure 3B).

The binding protein of ZMIZ1-AS1. (A) Silver staining following after RNA pull-down assay. (B) Western blot assay using hnRNPA 1 antibody
4 DISCUSSIONGefitinib is a first-line treatment drug used to treat lung cancer with sensitive mutations, which can significantly improve their overall survival rate. However, acquired gefitinib-resistance is still a major problem in cancer treatment and limits the therapeutic effect. At present, the underlying mechanisms of gefitinib resistance are not thoroughly studied. At present, EGFR 20 exon T790M mutation, MET amplification and tumor cell phenotypic transformation are the most common causes of acquired drug resistance.7-9 However, the secondary drug resistance of tumors is complex, and there is still a lack of predictive indicators of curative effect and effective intervention strategies.10
Recent studies have found that ncRNA plays an important role in the development of drug resistance in tumor cells by controlling gene expression and translation at transcriptional or post-transcriptional levels. In our study, lncRNA ZMIZ1-AS1 was screened and identified from the microarray data in the GEO datasets. Consistent with results in the microarray data, ZMIZ1-AS1 was expressed at a higher level in gefitinib-resistant cells compared with gefitinib-sensitive cells. ZMIZ1-AS1 knockdown could significantly improve the sensitivity to gefitinib in gefitinib-resistant cells. The results demonstrated that ZMIZ1-AS1 can promote gefitinib resistance in NSCLC.
Growing evidence has pointed to the notion that many lncRNAs can function to regulate some target gene expression through direct interaction with proteins. hnRNPA1 is the most abundant and ubiquitously expressed member of heterogeneous nuclear ribonucleoproteins (hnRNPs). It is a RNA-binding protein associated with complex and diverse biological processes such as processing heterogeneous nuclear RNAs into mature mRNAs, RNA splicing, transactivation of gene expression, and modulation of protein translation.11-13 hnRNPA1 has been reported to participate in multiple molecular events in cancer transformation.14-16 Here, we reported that ZMIZ1-AS1 binded to hnRNPA1 by RNA pull-down assay and mass spectrometry. Our study has limitations because we did not reveal how ZMIZ1-AS1 promoted gefitinib resistance by combining with hnRNPA1. The specific mechanism needs further study.
In summary, the present study showed that ZMIZ1-AS1 promoted gefitinib resistance by binding with hnRNPA1. These findings enhanced our understanding of ZMIZ1-AS1in NSCLC gefitinib resistance. So far, there has been no report on the involvement of ZMIZ1-AS1 in the process and molecular mechanism of gefitinib resistance in NSCLC, so this study is original and innovative.
ACKNOWLEDGMENTThis work was supported by scientific research project of Jiangsu Commission of Health (M2020032).
CONFLICT OF INTERESTThe authors declare that there is no conflict of interests.
AUTHOR CONTRIBUTIONSChangwen Jing wrote the manuscript and designed the study. Haixia Cao and Rong Ma performed the real-time PCR experiments. Jianzhong Wu was responsible for cell viability and flow cytometry assays. Junying Zhang and Siwen Liu performed RNA pull-down analysis. Zhuo Wang contributed to the design of the study. All authors read and approved the final version of the manuscript.
ETHICS STATEMENTThe research didn't involve human ethical issues.
BiographiesChangwen Jing, an assistant researcher in clinical cancer research center of Jiangsu cancer hospital, has been focusing on the molecular mechanism of lung cancer.
Haixia Cao, a researcher in clinical cancer research center of Jiangsu cancer hospital, has been focusing on cancer biology.
Jianzhong Wu, the head of clinical cancer research center of Jiangsu cancer hospital.
Rong Ma, a researcher in clinical cancer research center of Jiangsu cancer hospital, has been studying on cancer research.
Junying Zhang, an assistant researcher in clinical cancer research center of Jiangsu cancer hospital, has been working on tumor research.
Siwen Liu, an assistant researcher in clinical cancer research center of Jiangsu cancer hospital, has been working on tumor research.
Zhuo Wang, an assistant researcher in clinical cancer research center of Jiangsu cancer hospital, has been focusing on tumor biology.
REFERENCES
1Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020; 70(1): 7- 30. 2Yamaoka T, Ohba M, Ohmori T. Molecular-targeted therapies for epidermal growth factor receptor and its resistance mechanisms. Int J Mol Sci. 2017; 18(11): 2420. 3Shen P, Cheng Y. Long noncoding RNA lncARSR confers resistance to adriamycin and promotes osteosarcoma progression. Cell Death Dis. 2020; 11(5): 362. 4Lu Y, Zhao X, Liu Q, et al. lncRNA MIR100HG-derived miR-100 and miR-125b mediate cetuximab resistance via Wnt/beta-catenin signaling. Nat Med. 2017; 23(11): 1331- 1341. 5Jing C, Cao H, Qin X, et al. Exosome-mediated gefitinib resistance in lung cancer HCC827 cells via delivery of miR-21. Oncol Lett. 2018; 15(6): 9811- 9817. 6Hu Y, Qin X, Yan D, et al. Genome-wide profiling of micro-RNA expression in gefitinib-resistant human lung adenocarcinoma using microarray for the identification of miR-149-5p modulation. Tumour Biol J Int Soc Oncodevelop Biol Med. 2017; 39(3): 1010428317691659. 7Balak MN, Gong Y, Riely GJ, et al. Novel D761Y and common secondary T790M mutations in epidermal growth factor receptor-mutant lung adenocarcinomas with acquired resistance to kinase inhibitors. Clin Cancer Res Off J Am Assoc Cancer Res. 2006; 12(21): 6494- 6501. 8Engelman JA, Zejnullahu K, Mitsudomi T, et al. MET amplification leads to gefitinib resistance in lung cancer by activating ERBB3 signaling. Science. 2007; 316(5827): 1039- 1043. 9Oser MG, Niederst MJ, Sequist LV, Engelman JA. Transformation from non-small-cell lung cancer to small-cell lung cancer: molecular drivers and cells of origin. Lancet Oncol. 2015; 16(4): e165- e172. 10Wu SG, Shih JY. Management of acquired resistance to EGFR TKI-targeted therapy in advanced non-small cell lung cancer. Mol Cancer. 2018; 17(1): 38. 11Jean-Philippe J, Paz S, Caputi M. hnRNP A1: the Swiss army knife of gene expression. Int J Mol Sci. 2013; 14(9): 18999- 19024. 12Roy R, Durie D, Li H, et al. hnRNPA1 couples nuclear export and translation of specific mRNAs downstream of FGF-2/S6K2 signalling. Nucleic Acids Res. 2014; 42(20): 12483- 12497. 13Guil S, Long JC, Caceres JF. hnRNP A1 relocalization to the stress granules reflects a role in the stress response. Mol Cell Biol. 2006; 26(15): 5744- 5758. 14Pino I, Pio R, Toledo G, et al. Altered patterns of expression of members of the heterogeneous nuclear ribonucleoprotein (hnRNP) family in lung cancer. Lung Cancer. 2003; 41(2): 131- 143. 15Li WJ, He YH, Yang JJ, et al. Profiling PRMT methylome reveals roles of hnRNPA1 arginine methylation in RNA splicing and cell growth. Nat Commun. 2021; 12(1): 1946. 16Zhu HE, Li T, Shi S, Chen DX, Chen W, Chen H. ESCO2 promotes lung adenocarcinoma progression by regulating hnRNPA1 acetylation. J Exp Clin Cancer Res. 2021; 40(1): 64.
留言 (0)