
{kind=link}
記住我
A common denominator marks the current literature on the analysis of animal and human movement-based behavior: on the one hand, the community is “brimming with excitement and boundless opportunity” being overwhelmed by the big kinematic data offered by novel computational technologies of machine vision and deep learning (von Ziegler et al., 2021). On the other hand, it witnesses a plethora of novel phenotyping methods of automated behavior classification and discovery, based on novel technologies borrowed from machine vision and feature extraction (Berman et al., 2014; Egnor and Branson, 2016; Calhoun and Murthy, 2017). Perhaps because in the vast majority of these studies the main interest is in the processes that mediate behavior, rather than in the growth and form of behavior in and for itself (as was the case with comparative anatomy before the discovery of genetics and neurophysiology), automated behavior classification and discovery largely focus on species-specific behavior, disregarding the phyletic and cross-phyletic perspective.
This essay offers a solution to the situation by describing common primitives and modules, by describing the organization of movement in vertebrates and arthropods, and by providing a comprehensive methodology for revealing and confirming this organization.
The methodology we offer presents a rebirth of several old, yet radical insights largely ignored in the study of movement-based behavior. The first, organismic or holistic insight is that the organization, including the primitives (natural elementary building blocks), modules (locally integrated units) and generative rules of the different levels of the pyramid of life cannot be defined separately and independently of the organism as an indivisible whole, without initial reference to the organization at other levels, and to the organism’s situatedness in the environment during all its developmental stages. The second, ethological, insight is that behavior is an extension of anatomy. It has been conceived by classical ethologists, hardly pondered upon, and abandoned. One implication of this insight is that the connectedness of the skeleton, with its specific mechanical constraints, is mirrored in movement. The other insights are structural. They have been central to developmental biology and evolutionary developmental biology (dubbed evo-devo), but shunned by ethologists, behavioral neuroscientists, and current day phenotypers of movement-based behavior. One such insight is that portraying growth and differentiation (morphogenesis) is essential for understanding the process through which a single-celled egg gives rise to a complex, multi-celled sentient, cognizing, and functioning organism. Another, continuity insight, is that “an organism’s skeleton begins as a continuum, and a continuum it remains all lifelong” (d'Arcy, 1942; Gould, 1971). Movement grows and differentiates continuously, much like living tissue, and not as a sequence of discrete, fragmented, units typically reported by classical ethologists and computational phenotypers (e.g., Berman et al., 2014; Datta et al., 2019; Ligon et al., 2019). Still another, connectedness insight is that a part of the body of an organism belonging to one species is homologous to a part belonging to another organism of another species, if and only if their connectedness to other parts of their respective bodies is identical (as opposed to the two parts having a similar shape). We use this so-called principle of connections, implemented by Saint Hilaire for the identification of anatomical homologies (Saint-Hilaire, 1830), for the definition of movement-based behavioral homologies.
Ethologists, behavioral neuroscientists and phenotypers either ignore, or take lightly, or adopt the opposite, reductionist views in the study of animal movement. Because of the above lacunae, the recording, presentation, analysis, and understanding of movement, being the primary component of behavior, has lagged behind the study of the corresponding molecular, genetic, and anatomical levels, for well over 50 years. The present work aims to fill these lacunae by creating a framework for implementing the holistic, morphogenetic, continuity, and connectedness principles in the study of the structure (geometry) of movement-based behavior.
To introduce the morphogenetic study of behavior we start with the primitives, modules, and geometric rules that have been isolated by evo-devo in vertebrates and in arthropods, and applied at the genetic and anatomical levels (Figure 1 top). Then we proceed to describe their counterparts in the study of movement. The primitives at the genetic level are genes, and their respective modules are gene regulatory networks (Wagner, 2007). The primitives at the skeletal level are bones in vertebrates and rigid exoskeleton segments in arthropods, and their respective modules are organs and the main axes of the body.
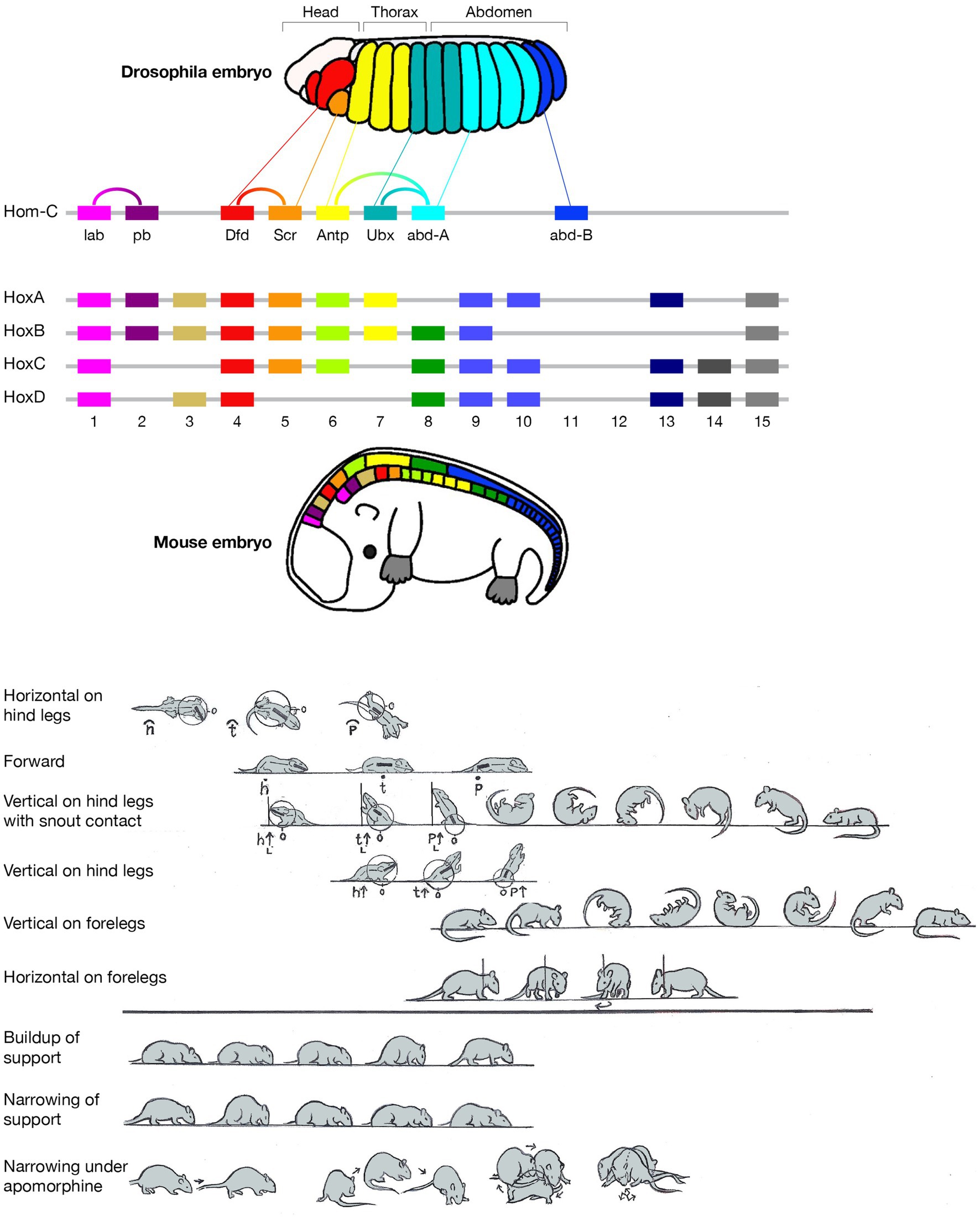
Figure 1. Schematic pictorial summary of the vertebrate bauplan: Top: The linear Hox genes’ organization on the chromosome(s) corresponds to their linear, modular, transcriptional expression along the Anterior–Posterior (AP) axis in an arthropod (fruit fly embrio) and a tetrapod (mouse embrio) (Adapted from Stefanie D. Hueber, Georg F. Weiller, Michael A. Djordjevic, Tancred Frickey, https://commons.wikimedia.org/wiki/File:Genes_hox.jpeg). consolidating into a kinematic linkage of multiple, specialized, rigid segments, bottom. This so-called collinearity between genes and the modular anatomical morphogenesis of the body plan is also manifested in a locomotor bauplan, unfolding on the temporal scales of moment-to-moment behavior (actualgenesis), ontogenesis, and partly also vertebrate phylogenesis. In vertebrates, locomotor behavior unfolds along six modular spatial dimensions, emerging successively in a fixed order and added progressively to the animal’s repertoire. The build-up is linear within each module separately, progressing from fore- to hind quarters. Pivoting and rearing on the hind legs emerges early in locomotor morphogenesis (top 4 horizontal lines) and pivoting and rearing on the forelegs emerges late (5th and 6th horizontal lines). In stressful situations and under the influence of dopaminergic drugs the spatial locomotor modules are eliminated from the animal’s repertoire in reverse order to their emergence, “last in first out.” Also, the support module is inactivated in a linear tail-to-head order (PA), restricting kinematic freedom.
Evo-devo shows that the morphogenesis of skeletal anatomy unfolds in reference to the main, Antero-Posterior (AP), Dorsi-Ventral (DV), and Proximo-Distal (PD) axes of the body. These axes are not extrinsic, arbitrarily superimposed geometric constructs, but rather real, active, partly independent physiological morphogenetic fields, now termed modules (Child, 1941; Opitz, 1985; Wagner et al., 2007). The demarcation of the anatomical body plan’s main axes is one of evo-devo’s ways to impregnate, right from the start, the primitives, modules, and generative rules of morphogenesis with holistic features even when dealing with local processes. The term body plan refers to a set of morphological features characterizing the members of a phylum of animals.
In our studies of animal movement, the primitives, modules and architectural plan of movement are induced by the anatomical body plan. Therefore, they critically mirror the mechanical connectedness imposed by the kinematic linkage of segments. The primitives are circular movements of single rigid segments moving within individual spheres schematically centered on the respective joint closer along the linkage to the base of support. A change in the base of support may entail a shift of the sphere’s center to the opposite joint, reflecting a respective reversal in the mechanical hierarchy between the segments. The coordinate systems of the individual spheres are gyroscopically oriented in reference to the absolute frame. Describing movement in reference to both the base of support and the absolute physical environment secures a relevant, situated (being embedded in the environment), and being embedded in the body description.
As in evo-devo, isolating primitives that are defined by their connectedness to other primitives, and by their relative position within the whole kinematic linkage, as well as by their orientation in reference to gravity and to absolute and body-related spatial direction, secures the global coherence of behavior, yielding natural kind (Wagner, 1996; Stanford Encyclopedia of Philosophy, 2022) primitives, modules, and generative rules, all permeated from the start with organismic features.
In contrast, classical ethologists and current computational phenotypers of behavior typically employ a reductionist approach. They first establish elementary building blocks, focusing exclusively on their intrinsic features and ignoring their extrinsic connectedness, and their being parts of an indivisible whole, be these units behavior patterns (Tinbergen and Perdeck, 1950; Tinbergen, 2020), stereotyped actions (e.g., Berman et al., 2014), sub-second modules (e.g., Wiltschko et al., 2015), “expert defined categories” (Anderson and Perona, 2014; Egnor and Branson, 2016), “postures” (Schwarz et al., 2015), or “discrete behavioral events” (Golani, 1973; my own youthful folly). Only then do they attempt to relate these units to each other using, e.g., motivational, statistical, or cluster analysis models. This procedure sidesteps the basic zoological procedure of relating, measuring, and comparing identically connected (homological; see below) and organismic (body plan) relevant structures. Assuming initial, seemingly unbiased lack of knowledge, they end up with a list of fragmented, hardly-related and non-generalizable categories that could not be assembled into a unified architectural plan nor could they be compared, except for relative frequencies, across treatments, or taxa.
The current essay singles out two geometric properties - linearity and modularity - and highlights their manifestation across the genetic, anatomical, and behavioral levels, built on studies in evo-devo and animal movement-based behavior (Figure 1 Top). A linear topographical order of the Hox toolkit genes on the chromosome(s); a collinear order of their expression; serially connected anatomical modular segments linearly ordered along the Antero-Posterior (AP) axis; their movement linearly recruited along the AP axis; separately and sequentially along each of six coordinative spatial modules in actualgenesis, ontogenesis, and partly also in phylogenesis, neurological recovery, adult intact and drug induced behavior, and agonistic interactional behavior.
The progressive growth and differentiation of movement within and across spatial modules enacts a buildup in the organism’s neurological freedom of operational space. An opposite narrowing of operational space is induced by stress and dopamine agonist drugs. The organism’s operational space, encompassing action, perception, attention and primordial cognition and emotion, mirrors the dimensionality of physical space, including the continuity within, and discontinuities across, physical dimensions, the constraints imposed by the lower-level anatomical body plan and the genetics supporting it, and the affordances offered by the animate environment.
This essay offers a new synthesis portraying the morphogenesis of an organism’s genes anatomy and behavior architectural plan (bauplan, Figure 1B, bottom) in a comparative generalizable frame. The Anglicized term “body plan” was preceded by the German term bauplan (plural, baupläne), used in biology and introduced by Woodger in 1945, which means ground plan, architectural plan, or structural plan (Woodger, 1945; Hall and Hall, 1999; Rieppel, 2006).
2 Collinearity, modularity, and homology in the genes-anatomy-and-behavior architectural plan of vertebrates and arthropodsThe order of the Hox genes along the chromosome(s) is the same as their order of expression in the embryo. These toolkit genes generate along the head-to-tail axis of arthropods and vertebrates a linear modular partitioning of the body into a kinematic linkage of multiple, specialized, rigid segments (metameres). Using a movement notation description that mirrors the mechanical kinematic constraints on this linkage, we propose, above the vertebrate anatomical body plan, a collinear and modular bauplan of movement-based behavior (Figure 1 bottom), comprised of six relatively independent spatial dimensions, or modules. Movement within each module unfolds in moment-to-moment morphogenesis (actualgenesis), and in ontogeny, in the same linear, head-to-tail order: the modules emerge, grow and differentiate in a fixed order, building up the freedom of movement of the linkage. Whole body movement around the hindquarters precedes movement around the forequarters. The same order applies to forward progression across vertebrate phylogeny. In stressful situations and under the influence of drugs, the spatial modules are eliminated following a “last in first out” rule, while a support module folds in an opposite, tail- to-head linear order, restricting the freedom of movement of the linkage. The same modules unfold and fold in the same order in arthropods.
Because each locomotor dimension involves the management of specific perceptual inputs relating to a specific physical dimension, the bauplan also manifests a hierarchy of attention and of primordial cognition. The momentary kinematic freedom of movement exercised by the homologous linkage discloses the animal’s understanding of the situation, communicating to the organism itself and to its phyletic congeners the emotion indicated by the affordances it offers, thus exhibiting the organism’s life-world (umwelt). The performance of novel movement types enables the performance of still unforeseen, more advanced types, liberating the organism from being distracted by external and internal stimuli, enriching its life world by endowing it with an increasing freedom to act with precision or not to act, on the basis of internal context. The portrayed bauplan may serve as a road map in studies of the homeotic, neural, and aspects of the cognitive levels of the pyramid of life.
A central theme of this work is the growth of behavior from its inception, at any stage of life: birth, exit from a den, courtship, hunting, and other instances that recur in natural behavior following a structured, well-defined choreography. The synthesis of the built-in rules, according to which many behaviors form during growth processes, has not been properly recognized. While pointed out in the ethological literature, it has not yet achieved recognition as a central theme of ethology. Possibly the most enigmatic example, yet outstandingly flamboyant, is the “war dance” of stoats (Mustela ermine) and ferrets (Mustela euro), involving stoting, logrolling in the air, zigzags, spins, loops, and very fast bounces. It is discussed in detail in the Discussion section. The geometric analysis of the rules of this buildup and subsequent shutdown across species, placing it in physical space, relating it to genetics, anatomy, and neurochemical modulation, within a biologically relevant comparative framework, and focusing on its growth and form is a central theme of this essay.
The first step of defining the primitives of movement-based behavior in terms of their topological relatedness to the whole is critical. The pyramid of hierarchical control of living matter is distinguished by an ascending order of levels, each level featuring its respective simplest element or primitive: atoms at the physical level, molecules at the biochemical, genes at the genetic, …, cells at the cellular, …, rigid segments at the anatomical, organs at the skeletal, etc. Remarkably, however, there is as yet no scientific consensus over what are the primitives of the next-level-up in the hierarchy, that of movement and of locomotor behavior. At the same time, while the anatomical body plan features highly in evo-devo, there are as yet no portrayed baupläne that encompass the genes, anatomy and behavior levels within the same frame for any phylum.
The aim of this essay is to identify both the primitives of movement, as well as the bauplan of the morphogenesis of movement-based behavior in vertebrates and arthropods. As illustrated below, for comparative anatomists it has been imperative to attend simultaneously to the primitives and the plan. Borges’ account of “Funes the memorious,” as related by a linguistic anthropologist (Chrisomalis, 2010) in an essay on the effect of written numeral systems on human cognition, illustrating the problem and the endeavor lying ahead of us:
“a person blessed or cursed with an apparently limitless memory… who told the author… that in 1886 he had invented an original system of numbering and that in a very few days he had gone beyond the 24,000 mark. He had not written it down, since anything he thought of once would never be lost to him. His first stimulus was, I think, his discomfort at the fact that the famous 33 gauchos of Uruguayan history should require two signs and two words, in place of a single word and a single sign. He then applied this absurd principle to the other numbers. In place of 7,013, he would say (for example) Maximo Perez; in place of 7014, The Railroad; other numbers were Luis Melian Lafinur, Olimar, sulphur, the reins, the whale, the gas, the caldron, Napoleon, Agustin de Vedia. In place of 500, he would say nine. Each word had a particular sign, a kind of mark; the last in the series were very complicated. I tried to explain to him that this rhapsody of incoherent terms was precisely the opposite of a system of numbers. I told him that saying 365 meant saying three hundreds, six tens, five ones, an analysis which is not found in the “numbers” The Negro Timoteo or meat blanket. Funes did not understand me or refused to understand me” (Borges, 1962).
Chrisomalis wryly points out that “whether we say 7,013 or Maximo Perez is not simply a stylistic choice” (Chrisomalis, 2010). Indeed, the invention of numeral systems (numeral literacy) opened up for human cognition an immense arithmetic manifold, revealing structure and arrangement such as odd and even numbers, prime numbers, zero, ordinal numbers denoting order, and cardinal numbers denoting quantity, distance and proximity between numerals, numbers “wrapping around” upon reaching a certain value – the modulus, and the arithmetic operations, including comparisons – all nonexistent in Funes’s epistemological particularistic world.
von Uexkull (von Uexküll, 1957) and later Gibson (2014) stated that organisms first project an operational meaning (affordance) on a perception, and then act on it in a way corresponding to that affordance. This dictum necessarily also applies to Funes and to literate arithmeticians: for Funes the one and only affordance of “Maximo Perez” and “The Railroad” is counting and then comparing the respective frequencies of occurrence of these “numbers” (and hence their conditional probability of appearance) in a sequence of similarly labeled numbers. In contrast, for a literate arithmetician, the affordances of the respective numeral counterparts of these numbers – 70/4 – include ordering the two numerals, comparing their sizes, calculating their distance from each other, as well as performing these and all the other arithmetic operations with all the other numerals across the whole arithmetic manifold. This, in a nutshell, is the difference between the use of disconnected (Funes type) building blocks, and the use of primitives which are a priori designed to be defined by their connectedness. This is also the difference between the building blocks listed in computationally derived ethograms that initially ignore extrinsic connections, and the primitives of movement expected in an “anatomical” paradigm of behavior. Funes and the computational phenotypers are either blind or disinterested in the affordances offered by the connectedness of the manifold generated by the respective primitives comprising it, be they numerals in arithmetic or movement primitives in behavior. But since there are no such thing as self-sustained facts floating in non-entity, because every distinction implies a universe of discourse (Whitehead, 2010a; Whitehead, 2010b), the primitives listed by Funes as well as by most current computational studies of behavioral phenotyping start by assuming a lack of knowledge about the connectedness of behavior, ending up with a fragmented particularistic landscape. In contrast, the paradigm employed in comparative anatomy and by the movement studies presented below defines and then isolates well-connected (i.e homologous) primitives, yielding a well-connected coherent manifold (i.e., a bauplan).
Notations representing holistic features and connectedness highlight natural primitives and baupläne. Numeral notation has been critical for exposing the structural manifold of arithmetics because of the connected nature of numerals and their indication of the infinite whole. Chemical notation has become practically indistinguishable from the theory of chemical bonding (Bawden, 2001; Brock, 2012) because the notation represents chemical connectedness of atoms in terms of (covalent and ionic) bonds, referring to the periodic table of elements, a tabular display of the chemical elements, which are arranged by atomic number, electron configuration, and recurring chemical properties. The periodic table, together with chemical notation, provides a bona fide example of a holistic, connected representation of an architectural plan, in this case of chemical primitives (atomic elements) and modules (molecules). In music notation notes relate to octaves and scales, thus manifesting a fixed relative position, and providing the foundation for music analysis and composition (Nash, 2015). In the same way, Eshkol-Wachman Movement Notation (EWMN, Eshkol and Wachman, 1958; Eshkol and Harries, 2001, 2004) features primitives consisting of single movements, each performed by a single rigid segment, characterized by its relative position along the skeletal linkage, and always defined in reference to its kinematic effect on that whole linkage. This connectedness endows the movement primitive with its homological identity. The notation’s spherical System of Reference (SoR) portrays not only already occupied spatial dimensions, but also the potential for evolutionary behavioral innovation. Another asset of this movement notation is the distinction it offers between continuous vs. discontinuous slabs of movement. These features are discussed in detail in later sections.
Behavior is an extension of anatomy. Konrad Lorenz, a founder of the discipline of ethology, accepted his share in the 1973 Nobel Prize in Physiology and Medicine with the reflection that his and ethology’s “most important contribution to science” has been the discovery “that the very same methods of comparison, the same concepts of analogy and homology, are as applicable to characteristics of behavior as they are to those of morphology” (Lorenz, 1974). In particular, to the same extent that anatomists use bones like humerus and radius, and skeletal organs like head or right forelimb, to establish the concept of skeletal homology, ethologists have aspired to isolate the particulate processes of behavior in order to establish behavioral homologs. Half a century later, however, Lorenz is mostly remembered for his extensive documentation of the “fixed action pattern,” which he presumed to be the elementary building block of behavior and a waypoint, not the goal, of the quest for behavioral homologies. The fixed action patterns (Barlow, 1996; Lorenz, 2012) and their contemporary descendants are as idiosyncratic as the terms used by “Funes the memorious,” disallowing geometric scaling and/or comparisons of behavior.
A substantiation of the Lorenz insight that behavior is an extension of anatomy, requiring the same methods of analysis, thus sends us to comparative anatomy in its current manifestations: developmental anatomy and developmental evolutionary biology (evo-devo). Unlike Borges’ hero, we promote generalizability by portraying here the natural geometric manifold that unfolds (and folds) in this process, revealing a universal architectural plan that encompasses genes, anatomy, and behavior, including perception, attention, cognition, and emotion.
The topological definition of anatomical homology. A straightforward example of the ontology that was used in comparative anatomy for several centuries is presented in a French anatomy book published in 1555 (Figure 2). The concept of homology is the cornerstone of comparative anatomy. Comparing the presumed right upper arms of two animals belonging to two different tetrapoda taxa, the one and only way to ascertain that one is indeed measuring the respective right upper arms, is by making sure that both segments are articulated on their respective proximal ends to the rest of the linkage through spherical joints (shoulders), and on their respective distal ends through hinge joints (elbows), which are in turn connected to two parallel bones (radius and ulna), which are in turn articulated on their respective distal ends through synovial joints (radiocarpal joints)…etc. Defining the two bones as upper right arms or humeri establishes them as identical (homologous), tacitly implying a (homologous) body plan. There can be no rigorous science of comparative anatomy without ascertaining that a humerus is indeed a humerus, whether it is embedded in the arm of a human, the wing of a bird, or the foreleg of a horse, regardless of its respective form and function in these different taxa. Its identity is secured by Saint Hilaire’s (Saint-Hilaire, 1822) principle of connections, stating that “the sole general principle one can apply is given by the position, the relations, and the dependencies of the parts, that is to say, by what I name and include under the term connections (in Russell, 1945; see also Beer, 1974).” Bones that occupy the same relative position in the respective skeletons of different taxa (and genes that occupy the same relative position in the gene sequence of different taxa), are defined as homologous.
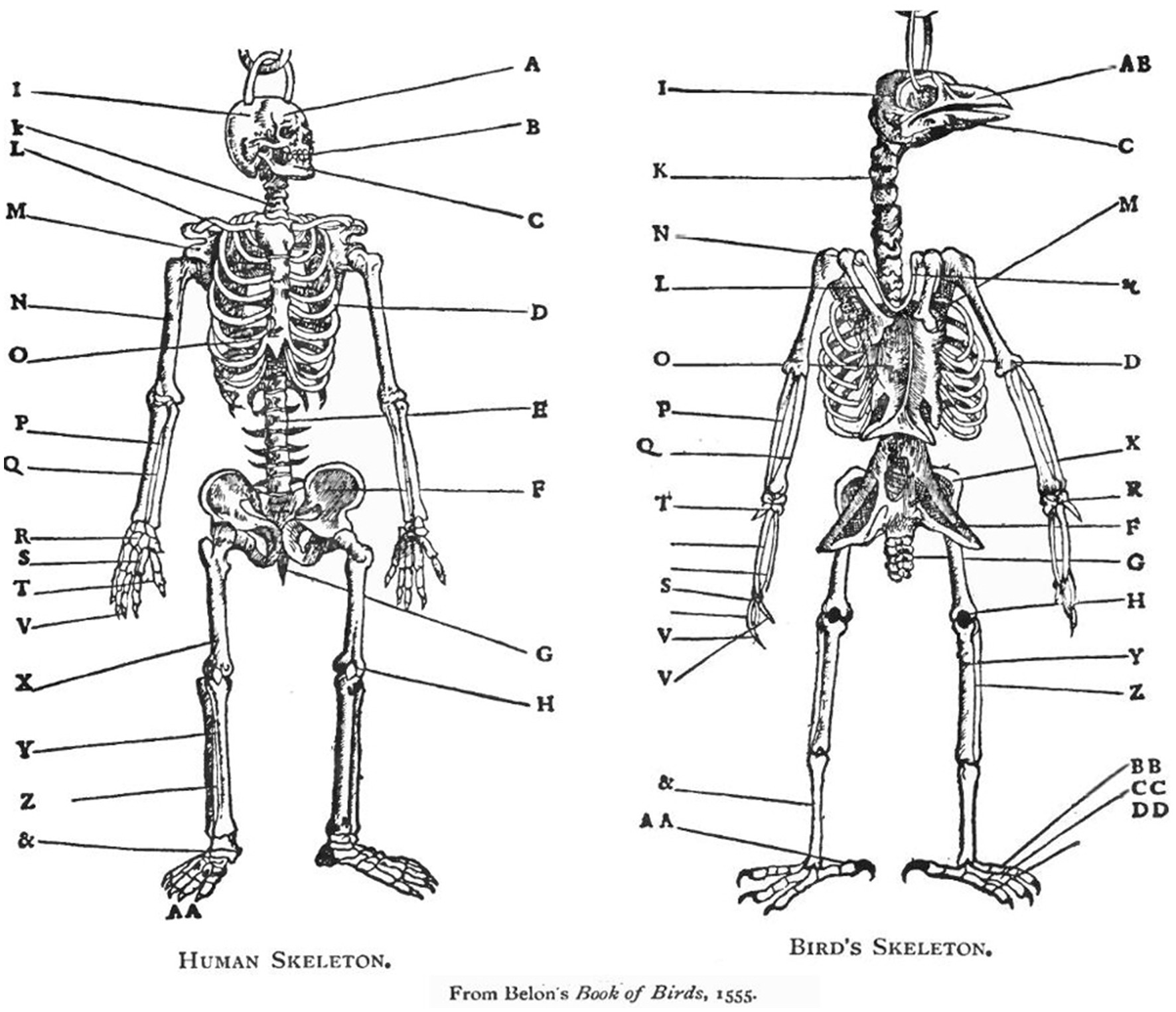
Figure 2. A human and a chicken skeleton share the same architectural plan (same body plan) (Belon, 1555). The whole skeletons, the respective organs (for example, the respective heads) as well as the individual bones (for example, the respective humeri) are homologous. According to the pre-Darwinian definition, the respective organs and bones are homologous because of their identical anatomical connectedness to other bones: they occupy the same relative position in the respective skeletons. According to the Darwinian definition they are homologous because they have descended from common ancestral bones and organs. Neither definition is based on the form or function of bones and organs. In the current essay we describe the homologous primitives (“bones”), modules (“organs”), and bauplan of behavior (Belon, 1555).
This topological definition of homology preceded and served as the basis for the later, Darwinian, definition of homology, which was based on common descent (Bateson, 1894). Needless to say, identity (homology) is currently secured by Darwin’s historical principle of common descent (Darwin, 1909). With behavior, however, historical evidence is absent. Therefore, the earlier, topological definition can still be useful if an invariant relative position of a behavioral primitive or a behavioral module can be demonstrated (Golani, 2012). The human and avian humeri are two manifestations of the same character identity (hence their sharing the same name), manifesting two distinct character states (Wagner, 2000), hence the differences between the humerus of a human and of a bird. Most important, the morphogenetic (developmental) history of the humeri is part and parcel of their definition (Hall, 1995). The use of the principle of connections in the definition of behavioral homologies is at the heart of the argument presented in this essay (for the primacy of a structural definition of homology; Wagner, 1994); for other definitions of homology (see Donoghue, 1992; Haszprunar, 1992).
Ethograms versus movement primitives. Having defined the presumed building blocks of movement on the basis of so-called expert decisions, or intrinsic features of the movements, or intrinsic features of the movement video-images, or the specific demands of a study, or unsupervised deep learning procedures (which not unlike Funes, rely on the cognizing or computational agent’s impeccable memory), provides a list of presumed building blocks, along with the probabilities of transitions between them. Take for example the list of “subsecond blocks of behavior” isolated in mouse open field behavior by an unsupervised deep learning procedure (Wiltschko et al., 2015). The automated procedure isolates body states from a 3D video, manifested as repeated pixel formations, and then counts their frequencies and reports the location and timing of their performance. The next stage includes manual labeling of the states. The labeling is independent of measurement and is based on subjective “expert evaluation,” not unlike classical ethologists observing behaviors and labeling them deliberately. For example, the deep learning program identified two states that were subsequently labelled “low rearing” and “high rearing.” However, this scaling of rearing episodes was subjective rather than data driven. Thus, even low and high rearing cannot be compared on any measurable scale, except for frequencies of performance, even in the same animal and in the same session, let alone in a comparison of “rearing” states across taxa.
In contrast, a description of the orientation of the parts of the trunk in reference to individual spherical coordinate systems schematically centered on their respective caudal joints would sensitize and prompt the observer to perceive and record the behaviors illustrated in Figure 3 as head raising (left), torso raising (middle) and pelvis raising (right) as three increasingly larger amplitude movements, along the same, vertical, dimension, recruiting an increasingly larger number of trunk segments, in an antero-posterior (AP) order: first the head, then the torso, which also carries along the head, and then the pelvis, which also carries along the torso and the head. In contrast to the fragmented list yielded by an ethogram, the kinematic primitives yield a fully connected description disclosing the kinematics of a linkage as well as the manifestation of a behavioral spatial module. The performance of these movements in sequence may constitute a morphogenetic buildup process, and the geometrical nature of this type of description allows a comparison across taxonomic groups. Unlike the building blocks of ethograms, the primitives illustrated in Figure 3 (i) refer to the whole kinematic linkage (much like the specific bones in a skeletal body plan), and (ii) can be assembled into higher level modules (here a vertical spatial module), which may amount to a whole body rotation around the side-to-side axis of the body of 360° amplitude (Figure 1, vertical on hind legs with snout contact), which can in turn be embedded within a whole body bauplan.
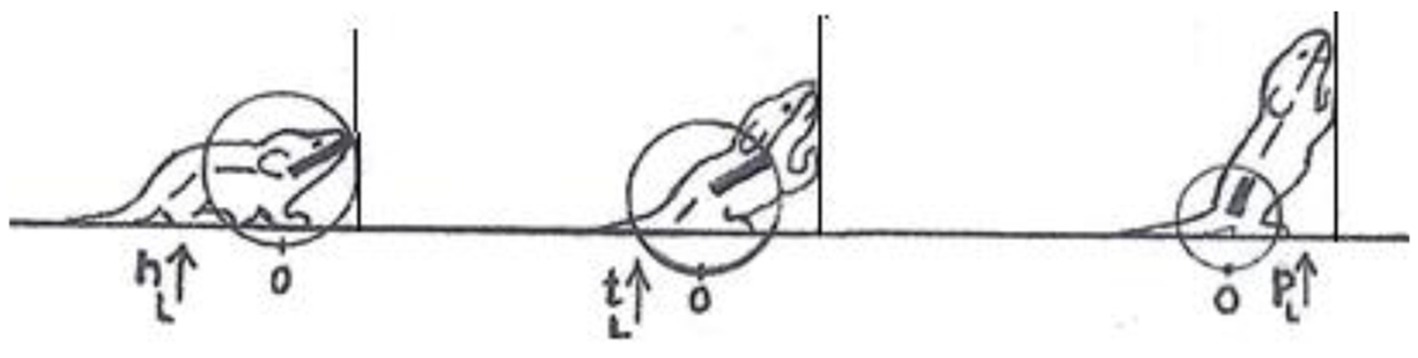
Figure 3. An illustration of three primitives of movement in an infant rat (Rattus norvegicus): head vertical movement (left), torso vertical movement (middle), and pelvis vertical movement (right). The straight lines represent the parts of the trunk. The circles stand for the spherical coordinate systems, respectively, centered at the root of the movement. The heavy lines stand for the respective part of the trunk that moved. During head movement (left) only the head changed its angular relation to its next caudal part; during torso movement (middle) only the torso changed its angular relation to its next caudal part, carrying along the neck and the head, and during pelvis movement (right) only the pelvis moved carrying along the torso, the neck and the head. Each of the notational expressions written under the illustrated movements stands for a primitive movement-type. The 3 illustrated movements constitute the beginning of the fourth natural module illustrated in Figure 1 as the “vertical on hind legs with snout contact,” which amounts to tumbling pelvis-on - a backwards whole-body rotation around the side-to-side axis of the body (Modified from Eilam and Golani, 1988).
Segmentation of behavior based solely on deep learning cannot yield a coherent model of the organization of behavior. Using it without impregnating the presumed building blocks with extrinsic connectedness illustrates “the fallacy of the first step,” which says that “climbing a tree is not the first step to the moon” (Bar Hillel in Dreyfus, 2012). In contrast, a literate kinematic description has the potential of demonstrating the connectedness and generalizability of movement-based behavior. Since the obtained description is morphogenetic, it furthermore, specifies generative rules rather than conditional probabilities.
To flesh out Lorenz’s insight, the movement primitives of vertebrates and arthropods should yield a bauplan: an overall natural geometrical manifold of behavior consisting in natural kinds (Wagner, 1996; Stanford Encyclopedia of Philosophy, 2022; Bird, 2018) in the same way that in comparative anatomy bones form organs, which form a body plan. The homology of the primitives we seek to define (single movements of single rigid segments) is established by (i) demonstrating their connectedness (Saint-Hilaire, 1822) to other geometrically-defined single movements and by (ii) embodying in their definition the indivisibility of the growing and differentiating organism.
To sum up, the major fallacy shared by Lorenz and by ethogram constructors of all times, including the current computational phenotypers of behavior, is their failure to even recognize the need for primitives that derive their identity from their relative position in the bauplan, a practice which is taken for granted and implemented en passant by anatomists. The very labeling of a bone as a humerus serially implies the whole body plan. In this way each and every skeletal primitive becomes a topologically invariant, and therefore generalizable and comparable unit. In the same way, whereas “scanning” is a particularistic mnemonic label for a behavior, a “positive vertical head movement “is a topologically invariant primitive because its first performance following extended immobility always precedes the first positive vertical chest movement, and always follows the first forward trunk movement. Because of its fixed relative position in the sequence it is comparable-across-phyla.
The use of ethogram-type, essentially disconnected, behavioral building blocks and categories, whether defined on the basis of their intrinsic properties, or ad hoc, has been useful in fields such as behavior genetics, behavioral pharmacology, and neuroethology. In these fields, the subject of interest is not the organization or structure or comparative study of behavior, but rather the study of the lower levels of the pyramid of life, such as the biochemical, physiological, genetic, and neural levels, whose connectedness, continuity, structure, etc., are the focus of interest. It is the connectedness and continuity in these underlying levels, which are both supplied and enriched by the quantification of behavioral markers and measures. In these disciplines, even crudely and intuitively defined behaviors can often be useful as a springboard for studying the levels that mediate behavior. Studies in these disciplines, however, largely do not contribute to the comparative study of the structure, organization, or connectedness of behavior itself. While quantification of homologous behavioral primitives is not an absolute necessity in these fields, phenotyping in terms of homologous natural kinds (Wagner, 1996; Stanford Encyclopedia of Philosophy, 2022; Bird, 2018) would be more likely to improve measurement (for the current feasibility of such endeavor, based on high throughput tracking, see section on “Key managed kinematic quantities and the organism’s umwelt”).
Anatomical modules. In comparative anatomy the bone primitives whose definition includes their morphogenetic history (Wagner, 1989; Hall, 1995) are embedded within organs, which are the modular ontological components of comparative anatomy. For example, a humerus, a radius, an ulna, metacarpals, and phalanges are the constituents of a forelimb. The definition of this module also encompasses its entire morphogenetic history, as a discrete, quasi-independent, locally integrated process and individualized yet interconnected part, or developmental field (Opitz, 1985; Shubin and Hall, 1994; Raff and Valentine, 1997). The right forelimb module, the head, the trunk, or the right hind leg, are not just products of a reasonable division of the organism’s body into parts; but, rather, biologically-real entities of a profound morphogenetic unity. As with the primitives so too with the modules: the embryological development of complex structures is determined and controlled in a spatially coordinated, temporally synchronous, and segmented hierarchical manner that on the one hand implies the whole organism, and is defined by its relative position (connections), and on the other hand expresses both non-specific (that is, phylogenetic, manifesting character identity) and species-specific genetically-coded developmental information (manifesting character states). To cite an example of this type of organization manifested in common anomalies: “identical anomalies produced by such different causes as the presence of an abnormal number of chromosomes in a cell, gene mutation, teratogenic chemicals, and certain surgical procedures show that embryonic primordia respond as units in the production of anomalies of anatomical structure. Hence, they must also act as units during normal ontogeny. The presence of identical malformations in different mammalian species identifies developmental and anatomical homology. These abnormal and normal morphogenetic reactive units are the equivalents of the classical experimental embryologist’s regeneration fields, which are those units of the embryo in which the development of complex structures appropriate to the species is determined and controlled in a spatially coordinated, temporally synchronous, and involving a conservation of form in a hierarchical manner that expresses both phylogenetic and species- specific genetically coded developmental information” (Opitz, 1985).
What is to be expected of movement modules. To flesh out Lorenz’s insight further we show below that, in the same way that bone primitives are organized into organ modules, distinct chords of single movement primitives can be organized in kinematic modules. These modules have a distinct morphogenetic history, and are performed in a relatively discrete, quasi-independent, locally-integrated process along distinct spatial dimensions. For example, horizontal movements that start at the head, and grow in amplitude to progressively recruit all the parts of the trunk in whole-body horizontal movement (whole-body pivoting around the hind legs; Figure 1 bottom, top line) behave as a distinct physiological subsystem. But “horizontal” tacitly implies “vertical,” even as a future morphogenetic option or absence thereof, and implies a dimensional operational space, even as an operational option. Anatomical modules (organs) are relatively few in kind and number, with a lot of empty phenotypic space between them (Alberch, 1989), e.g., there is no organ that consists in a combination of a head and a forelimb. If indeed behavior is an extension of anatomy, we would expect that, at least during the early stages of behavioral morphogenesis, the locomotor modules will also be distinct, their number relatively small, and they will be added to the organism’s movement repertoire separately and sequentially, implying a separate “bookkeeping,” module by module.
Body plans. The body plan concept has formed the backbone upon which much of the developmental and evo-devo current research is anchored (Willmore, 2012). Remarkably, the 35 body plans characterizing the currently extant 35 phyla of multicellular organisms all appeared about 550 million years ago, when fossils first appeared (Conway Morris, 2006) during an apparent, relatively sudden “explosion” (Raff, 1996; Marshall, 2006). No new body plans have emerged since (Fitch and Sudhaus, 2002), nor have any combinations between body plans been formed, suggesting that each of these 35 architectures manifests a solution whose intact entirety is critical for life (Riedl, 1978; Wagner and Laubichler, 2004). Multicellular organisms are composed of multiple modules, with each module characterized by its distinct morphogenetic history and its final shape. All the modules, together with their characteristic invariant connectedness, form the organism’s body plan (Woodger, 1945; Eldredge, 1989; Raff and Valentine, 1997).
The central role played by the body plan concept is exemplified in the development of the serially repeated set of vertebral structures and somite in vertebrates (Raff, 1996), or in the development of the limbs (Shubin et al., 1997; Jeffery et al., 2018) or in the morphogenesis of rhombomeres in the transiently dividing segments of the developing neural tube within the hindbrain region (Krumlauf, 2016). In both arthropods and vertebrates the trunk is partitioned into specialized groupings of modules like the cephalon, thorax, and pygidium (in trilobites); the head, thorax, and abdomen (in insects); or the head, neck, upper and lower torso, and pelvis (in Tetrapoda). Laterally disposed, equivalent body parts (antimeres), are articulated on both sides of the anteroposterior (AP) axis, like fore- and hind limbs in vertebrates and paired appendages in arthropods, themselves segmented along the Proximo-Distal dimension (Wagner, 1996; Raff and Valentine, 1997). The body plan specifies the identity and the topology of the phenotype, without which the molecular mechanisms that fix, mediate, and support these features could not have been discovered, in the same way that the genetic basis of any complex phenotypic trait could not have been established without the awareness of this trait.
Since the main goal of the present essay is to also encompass behavior within the same plan, and the term body plan refers only to anatomy, we adhere here to the original term bauplan when also referring to behavior. If indeed behavior is an extension of anatomy, then each body plan should enact a corresponding behavioral bauplan. As with anatomy, morphogenesis must be the key for deciphering the organization of this bauplan, in the same manner that it is the key for deciphering the organization of an organism’s anatomical body plan. Since morphogenesis involves differentiation, the more general anatomical taxonomic characters and anatomical structures should appear in development before the more specific and the more specialized ones, as was indeed established for anatomy two centuries ago (Von Baer, 1828). Hence the primacy of a definition of body plans on the basis of their embryological history. The same principle should apply to behavioral baupläne.
As with anatomy, the bauplan of behavior is not only of significance in and of itself, but is also indispensable as a roadmap for exposing the molecular mechanisms that mediate it.
What is to be expected of movement baupläne? Having defined bones as primitives, organs as modules, and the invariant topological connectedness of modules across development as an anatomical body plan, we may join a founder of the study of the embryology of behavior, George Ellet Coghill (Coghill, 1929) in his rhetorical question: “what is (skeletal) anatomy for?” with the obvious answer being “(skeletal) Anatomy is for behavior” (Herrick, 1941). Coghill noted in 1929 that “it is hard to understand why the same method which has proved so suggestive in comparative anatomy has not been pursued more vigorously by embryologists.” But whereas Coghill focused on behavior as an extension of neural anatomy, we focus on behavior as an extension of skeletal anatomy. Behavior, in the context of a kinematic linkage of rigid segments has a geometrical structure. Thus our subject matter is that of the natural geometry of behavior, a geometry that can in turn serve as an explanation for the supporting anatomy. The rest of this essay is dedicated to geometrical aspects of the genes, anatomy, and locomotor behavior bauplan. Locomotor behavior relates to the spatio-temporal forms “carved” by the articulated body in space and time.
The dimensionality of a bauplan. Back in 1918, the (mathematician and) custodian of biological form (d'Arcy, 1942) wrote a letter to the (mathematician and) custodian of “process philosophy” (Emmet, 1966):
“My dear Whitehead, I have been thinking, or dreaming, lately over a matter of which I know so little… the first difficulty seems to me, to decide whether there are, in reality, three dimensions of space; and the second question is, whether there be, in reality, three dimensions or no, how did we come to think, or to find out, that it is so.
I suggest, that we are … fundamentally guided, by the influence of Gravity…we are always face to face with a vertical axis, and with a plane (or apparently plane) surface perpendicular to it. In other words, the right angle assumes a very special importance, and, consciously or unconsciously, we refer everything in space to trihedral coordinates. Now suppose… that we were of so minute a size… that gravity would have no sensible hold upon us; and suppose, owing to our minute size, that we were mainly under the influence of other, say molecular, forces. Then, to begin with, we should know nothing about a vertical, and care nothing about a right angle. And suppose, in the next place, that we lived in some sort of “close-packed” or crystalline medium, say a tetrahedral one, we should never dream of three-dimensional space …So, paradox or no paradox, I seem to be driven to the conclusion that there is a quibble, or even a fallacy, underlying our definition of Space, or of Dimensions, (or perhaps both). Perhaps that dimensions are not necessarily rectangular: or that perpendicularity, inter se, is not a fair condition to postulate of them. That the Space which actually exists is quite independent of dimensions… does that in any way prove that we have a right to say there are, in reality, three dimensions; is it anything more than a mathematical figment, an elegant formula…. given a symmetrical individual in symmetrical space, how on earth could you ever teach him what right and left meant. He would obviously have no right and left and space itself has, obviously, no right and left. And so, I come back to my query. Has Space really three dimensions; or is this only a convenient figment of terrestrial, and large and clumsy, mathematicians?
Ever yours faithfully,
D’arcy Wentworth Thompson.”
Thompson, whose concern is the growth of biological form, appears to be consulting Whitehead (2010a, 2010b) about the ontological status of spatial dimensions: Does physical three-dimensional space exist in and of itself, regardless of an organism’s biological context? The current essay shows how, while the animal’s three-dimensional life-space is enacted dimension by dimension in morphogenesis, in reference to the physical vertical absolute (gravity), the life-space is embedded within physical space, enfolding its dimensionality, and thus disclosing the animal’s (cognitive) understanding of it.
To provide a context for the geometry of the animal’s operational world and the behavioral bauplan enacting it, we first portray below the geometry underlying the main morphogenetic processes leading to the construction of the animal’s anatomical body plan: collinearity between the toolkit (Hox) genes’ topographical alignment, Hox order of expression along the body, morphogen shaping of the AP axis, cell movement along the AP axis, and movement-generated axial forces exerted by the newly-formed muscles shaping the skeleton.
The Hox genes are aligned in a fixed order on the chromosome. Our geometrical journey starts with a small number of “master” genes assembled in one or several clusters on the chromosomfvs of arthropods and chordates, aligned in a fixed order on the chromosome (Gaunt, 2018). The Hox are transcription factor genes: Hox proteins encode and specify the characteristics of “position,” ensuring that the correct structures form in the correct places in the body. The expression of a Hox gene in a cell confers segmental or positional identity to that cell, but does not form the actual segment itself.
Collinearity between Hox genes’ physical alignment on the chromosome and their order of expression. In the second station of our geometrical journey, the Hox genes are expressed in the embryo’s body, along the AP axis, in partially overlapping domains, and in the same linear order of their alignment on the chromosome(s). The developmental characteristics of each zone reflect the combination of Hox genes that are activated or repressed in its cells. The geometrical correspondence between the physical alignment of the genes and their order of expression has been termed collinearity (Lewis, 1978, 1985; Lewis et al., 1980). Because Hox gene activation is concurrent with axial extension and limb growth, the different areas of the body and limbs express distinct combinations of Hox gene expression. During the early stages, Hox expressed and other morphogen concentration gradients span the entire length of the embryo. Their relative amounts signal, at precise AP and DV positions defined by their longitude, latitude and altitude coordinates, the topographical location of limb buds on the embryo’s envelope (Carroll, 1995). Which Hox gene turns on, where does it turn on, and when does it turn on is foundational in evo-devo (Goodman and Coughlin, 2000; Papageorgiou, 2021).
Morphogen and cell movement both shape and are shaped by the AP and DV axes. In the third station of our journey, dividing cells exhibit targeted movement to specific sites to form tissues and organs (Solnica-Krezel, 2005; Ananthakrishnan and Ehrlicher, 2007). Morphogen and cell movement both determine and are determined by the AP and DV axes. Morphogen gradients guide cell movement. In mice for example, cell movement specifies the AP body axis (Migeotte et al., 2010). Directed movement and other rearrangements that shape organs and body axes involve a fine-grained control of cell polarity. One type of cell polarity is established in reference to the axes of the embryo or organs (Veeman and McDonald, 2016). Vertebrate gastrulation involves four evolutionarily conserved morphogenetic movements governed by signaling pathways. Two of them, convergence and extension, are associated with axis elongation in both vertebrate and invertebrate embryos (Supplementary Video 1). The process, termed convergent extension, involves the lengthening and narrowing brought about by the collective heaping up of cells toward the embryonic axis (Wallingford et al., 2002). Cell movement related to the main axes is a substantial component of overall cell movement during certain stages of embryogenesis (Solnica-Krezel, 2005).
The Hox genes implement the principle of connections along the AP axis. In the fourth station of our journey we find the linearly arranged modular end products of the morphogenetic process. In vertebrates, the embryonic AP axis is segmented into a fixed species-specific number of somites which varies across species, ranging from as few as 32 in zebrafish to 35 in humans, and to more than 300 in some snakes.
The modular developmental genetics of morphological homologs. Pondering about the apparently loose relationship between morphological characters and their genetic basis (Hall, 1995; Butler and Saidel, 2000; Wilkins, 2002), Wagner (2007) proposes that it is the historical continuity of gene regulatory networks rather than the expression of individual homologous genes that underlies the homology of morphological characters. These modular networks, referred to by Wagner as “character identity networks,” enable the morphogenesis of morphological homologs.
The ancientness of the Hox genes and of extant body plans. The discoveries of the Hox genes’ structure and function are among the most important discoveries in biology in the last few decades. These genes and their respective signaling pathways are shared by most living phyla, and are implicated in sculpting the body plan. They have already functioned more than 500 million years ago, before the famous Cambrian Explosion that gave rise to the 35 extant animal phyla mentioned above. These genes, their order on the chromosome, and their ordered expression, were so crucial for survival that their anatomical and operational sequencing had been preserved throughout this enormous span of animal evolution (Carroll, 1995; Durston, 2012). If movement-based behavior is an extension of anatomy, its bauplan should also be ancient, and its morphogenesis too should be shaped by Hox genes.
Collinearity between AP formation of modular organs and their order of recruitment in movement. Cell movement in reference to the main axes culminates in the linear AP formation of modular organs along the whole skeletal body plan (which also constitutes a module). Remarkably, these modular organs begin to be recruited in vertebrates in movement in the same linear order, separately along each of the six spatial modular dimensions. But in both the vertebrate embryo and neonate, during the transition from organogenesis to movement, muscle-induced mechanical load is involved, via chemical cues, in chondrocyte proliferation, shaping of individual bones, regulating the 3D organization of skeletal elements, and many other aspects of skeletal differentiation and growth (reviewed in Felsenthal and Zelzer, 2017).
The AP midline axis is a morphogenetic field used as reference for the anatomical Body plan. Evo-devo and developmental anatomy support the ontological status of the arthropod and vertebrate midline. This midline is not an imaginary abstract sagittal plane dividing the right from the left half of the body. It is a biologically real developmental field of profound morphogenetic importance: a part of an embryo that reacts as a spatiotemporally coordinated unit to normal localized forces of organization and differentiation.
The morphogenetic field construct, finding its earliest expressions in the beginning of the previous century (Harrison, 1918; Weiss, 1926; Child, 1941) was enhanced by Opitz (1985) and expanded by evo-devo to the now established morphogenetic module (Wagner, 1996; Wagner et al., 2007). If behavior is an extension of anatomy, then it should necessarily be organized in reference to, and around this midline AP axis field, and reflect its linear geometry (Figure 4).
留言 (0)