
記住我




Scg3 belongs to the Granin family, which encompasses chromogranin A (CgA), chromogranin B (CgB), and secretogranin II-VIII (Scg2-8), alongside other related proteins such as 7B2, NESP55, proSAAS, and VGF [20, 21]. These members exhibit diverse roles in regulating secretory pathways within endocrine and neuronal cells, including the modulation of granule formation and peptide processing, as well as the regulation of hormone and growth factor release facilitated by postsecretory granule-derived peptides [21].
Function of Scg3The Scg3 gene (SCG3), located on human chromosome 15q21, consists of 12 exons. The 3366-bp Scg3 mRNA encodes a 468-amino-acid acidic secretory protein characterized by seven pairs of consecutive basic amino acids [20]. Scg3 primarily functions as a regulator for the biogenesis of secretory granules and the secretion of hormone peptides in endocrine and neuroendocrine cells [14]. The Scg3 molecule exhibits at least three structural domains: a cholesterol-binding domain located on the N-terminal side, a CgA-binding domain situated in the central region, and a domain responsible for binding to the carboxypeptidase E (CPE) on the C-terminal side [22,23,24]. In secretory granules of Scg3-expressing pituitary and pancreatic endocrine cells, specific binding facilitates the localization of chromogranin A (CgA) to Scg3, thereby aiding in the sorting and retention of CgA into secretory granules [22]. Additionally, Scg3 binds to CPE, which is tightly bound to secretory granule membranes (SGMs) in a cholesterol-dependent manner, forming dense core aggregates essential for secretory granule biogenesis [14]Notably, Scg3 does not appear crucial for hormone and neurotransmitter secretion, as evidenced by the normal phenotype observed in Scg3-/- mice [16]. Scg3 is up-regulated in various tumors, including neuroendocrine tumors, small-cell lung cancer, prostate cancer, and hepatocellular carcinoma [25,26,27,28]. However, the precise function of SCG3 in tumorigenesis remains unclear. The extracellular regulatory role of Scg3 was elucidated through the ligandomics technique, which involves high-throughput identification of whole cell ligands coupled with simultaneous quantification of binding or functional activity [29,30,31]. Scg3 is identified as a disease-associated angiogenic factor, exhibiting minimal binding and angiogenic activity in normal vessels. However, its binding and angiogenic activity are notably increased under pathological conditions, indicating a crucial role for Scg3 in vessels under such circumstances.
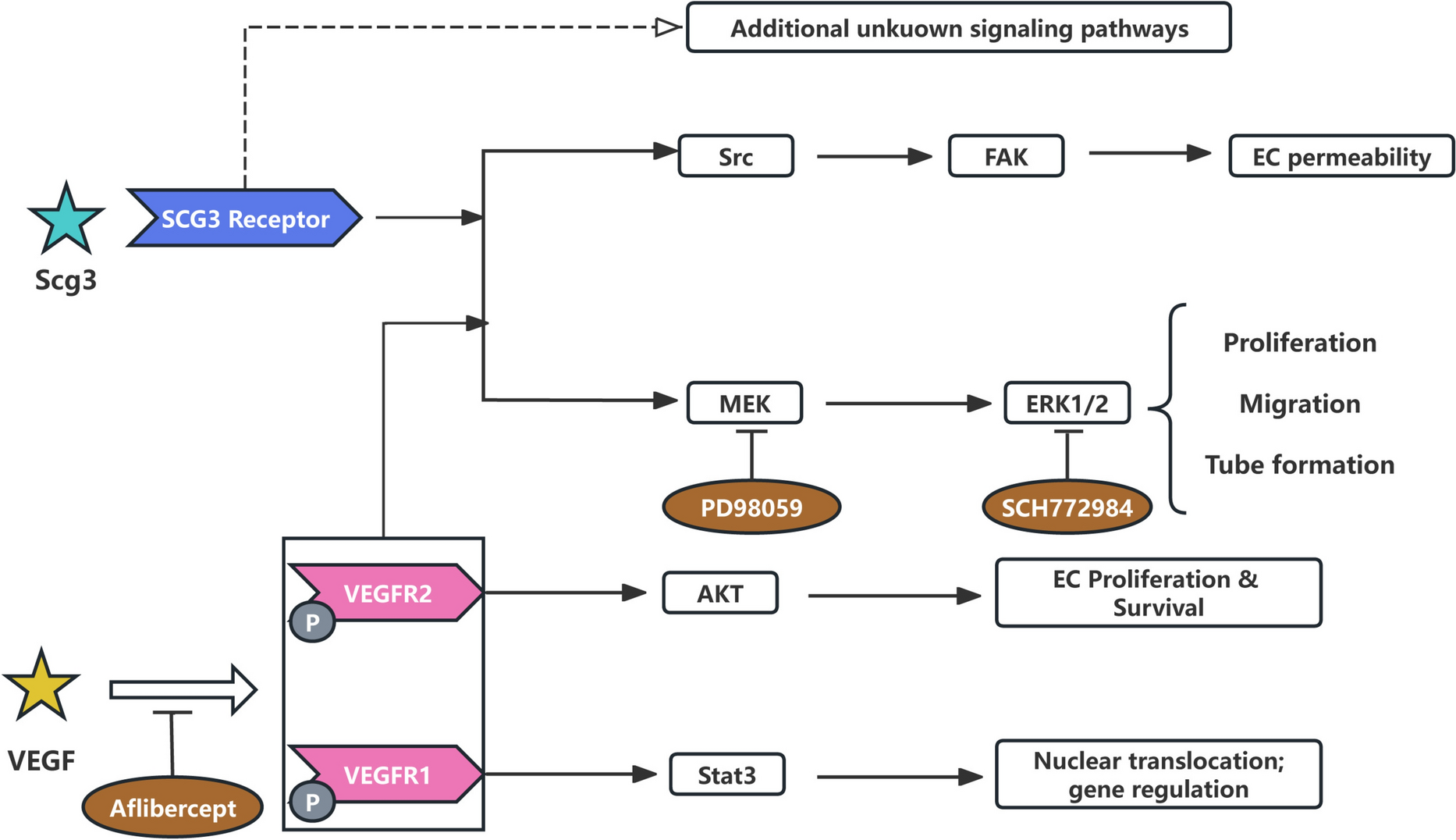
Molecular mechanisms of Scg3Scg3 signals through a non-VEGF-dependent receptor pathway (Fig. 1) [32]. Scg3 selectively promotes angiogenesis and retinal vascular leakage in diabetic mice but not in healthy mice. In contrast, VEGF induces both corneal neovascularization and retinal vascular leakage in both diabetic and normal mice [15]. Scg3 does not bind to VEGFR1 or VEGFR2. MEK/ERK can be activated by various receptors, including receptor tyrosine kinases (RTK), G protein-coupled receptors (GPCR), and integrins [33]. Hence, the Scg3 receptor may reside in RTK, GPCR, or integral proteins, providing a foundation for further Scg3 receptor identification. VEGF induces the phosphorylation of MEK, ERK1/2, Akt, Src, and Stat3, while Scg3 activates MEK, ERK1/2, and Src kinases, but not Akt or Stat3 [17]. Src, a non-receptor tyrosine kinase, plays a crucial role in the angiogenic signaling cascade and may be highly activated in pathological pre-retinal vascular clusters and ganglion cell layers [34]. These findings suggest that Scg3 and VEGF bind to different receptors and trigger distinct intracellular signaling pathways that converge on MEK/ERK and Src to regulate angiogenesis and vascular leakage. Inhibition of MEK using PD98059 blocks Scg3-induced proliferation, migration, and tube formation in human umbilical vein endothelial cells (HUVECs). Likewise, inhibition of ERK using SCH772984 suppresses Scg3-induced proliferation and migration of HUVECs, underscoring the pivotal role of the MEK/ERK signaling pathway in Scg3-induced angiogenesis [32].
Fig. 1
Scg3 molecular mechanisms. Scg3 does not binds to VEGFR1 or VEGFR2, Scg3 activates ERK and Src signaling pathways through unknown receptors, but not Akt and Stat3. ERK activated by Scg3 and VEGF can regulate various metabolic events. PD98059 and SCH772984 block signaling pathways by inhibiting MEK and ERK, respectively
Scg3 and intraocular neovascularizationScg3 is an intravesicular protein implicated in a diverse array of pathological processes, encompassing neurotransmitter storage and transport, secretory granule biogenesis, and regulation of angiogenesis [14, 15, 35]. Noteworthy, Scg3 exhibits strong associations with the progression of diabetic retinopathy, retinopathy of prematurity, wet age-related macular degeneration, and corneal neovascularization. Moreover, Scg3 has been identified as a more specific angiogenic factor [17,18,19]. The efficacy and safety of Scg3 inhibitors have been demonstrated in mouse models [36,37,38]. In summary, these studies collectively suggest that Scg3 represents a distinct neurogenic angiogenic factor.
Scg3 and diabetic retinopathyDR, a complication of diabetes, stands as the leading cause of vision loss among working-age adults[39]. Early diabetic retinopathy presents with endothelial cell (EC) and pericyte apoptosis, vascular leakage, and leukocyte adhesion, potentially progressing to microaneurysms, retinal vein occlusion, diabetic macular edema, and proliferative diabetic retinopathy [40]. Proliferative diabetic retinopathy (PDR) typically arises in the advanced stages of DR, characterized by fibrovascular proliferation leading to neovascularization, vitreous hemorrhage, and tractional retinal detachment [3, 41]. Beyond microvascular concerns, substantial evidence indicates that microvascular and neurodegenerative complications are integral components of DR pathogenesis [42].
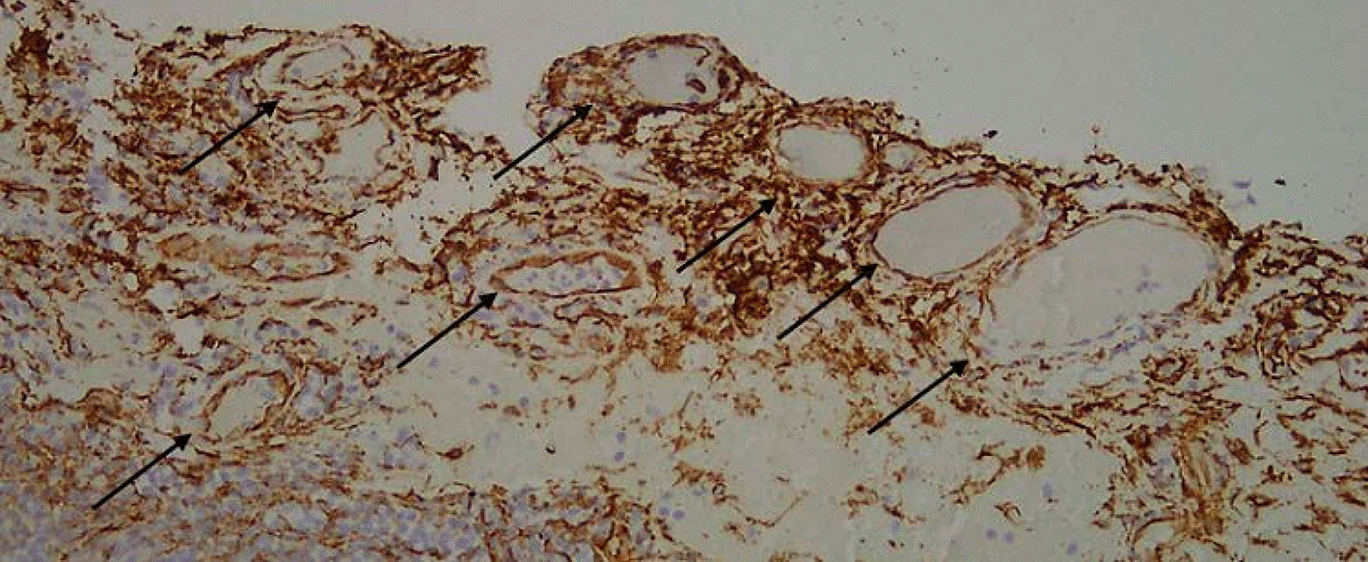
In 2017, LeBlanc et al. demonstrated significantly increased binding of Scg3 to diabetic retinal endothelial cells [17]. This finding implies that Scg3 may play a pivotal role in the development of retinal neovascularization under pathological conditions. Subsequent investigations revealed Scg3 expression in retinal neurons within the ganglion cell layer (GCL), inner plexiform layer (IPL), outer plexiform layer (OPL), inner photoreceptor segment (PIS), and retinal pigment epithelium, with limited expression in the inner nuclear layer (INL), outer nuclear layer (ONL), and outer photoreceptor segment (POS) [17]. Scg3 was found to be secreted from dysfunctional β cells, suggesting potential upregulation in type 1 diabetes [44]. Moreover, platelets in atherosclerotic lesions release Scg3, contributing to vascular complications associated with diabetes [45]. In diabetic mice, Scg3 expression in retinal vessels showed a modest increase of 1.38-fold. On the other hand, the surface binding of Scg3 in retinal vessels of diabetic mice exhibited a remarkable 1731-fold increase, indicating a significant upregulation of Scg3 receptors in diabetic vessels. Furthermore, Scg3 selectively promotes vascular leakage from the deep vascular plexus in diabetic retinopathy. Additionally, Scg3 selectively promotes vascular leakage from the deep vascular plexus in diabetic retinopathy. Ji et al. employed a unique in vivo ligand-binding assay, revealing a 22.7-fold increase in Scg3 binding to the retinal vasculature of diabetic mice compared to that of healthy mice. CD31 (PECAM-1), a well-known endothelial marker and endothelial junction protein, demonstrated predominant binding of Scg3 to the CD31-deep retinal vascular plexus under diabetic stress conditions, contrasting with relatively low binding to the CD31 + superficial and intermediate plexuses within the same diabetic retina[43]. In vivo functional analyses have independently confirmed that Scg3 specifically promotes angiogenesis and vascular leakage in diabetic mouse vessels, without affecting healthy ones [17, 44]. This suggests that Scg3 is a distinctive neurogenic factor that selectively impacts vascular and angiogenic processes in diabetes, leading to new interactions between the nervous system and blood vessels with a high level of disease specificity.
Scg3 and retinopathy of prematurityAs nerves develop, the retina undergoes vascularization, culminating in the formation of a functional neurovascular network. Impaired neurovascular coordination has been implicated in the development of ROP. While abnormal retinal vasculature is a hallmark of ROP, accumulating evidence suggests that the neural retina plays a significant role in the disease’s pathogenesis[45,46,47,48].
Contrary to expectations, Scg3 expression was not elevated in oxygen-induced retinopathy (OIR) compared to healthy retinas [17, 18]. Recently, Dai et al. employed a unique in vivo endothelial ligand-binding assay and quantification, revealing that Scg3 binds significantly more than sevenfold to retinal vessels in OIR mice [18]. Scg3 induces extracellular migration of secreted ligands that regulate both diseased and healthy endothelial cells. Angiogenic receptors on diseased cells are elevated and remain dormant until disease is present. It is important to note that Scg3 does not regulate physiological retinal neovascularization.
Knockout Scg3 mice (Scg3-KO) effectively reduced pathological retinal neovascularization in OIR. Scg3-KO rats showed no notable abnormalities in viability, fertility, or motor behavior. Additionally, Scg3-KO mice maintained a typical retinal vascular phenotype without affecting the developmental rate or structure of retinal vascular networks in all layers [19]. These findings highlight Scg3 as a distinctive angiogenic factor in neurogenic retinopathy of prematurity, demonstrating a significant level of disease specificity.
Scg3 and choroidal neovascularizationAMD is a disease in which the structure and function of the macula undergoes aging changes [49]. In advanced AMD, the clinical manifestations are dominated by atrophy and thinning of the outer retina, loss of RPE, and macular neovascularization (MNV). Based on the anatomical location of neovascularization and vascular composition determined by optical coherence tomography (OCT), MNV is classified into three subtypes: type 1, type 2, and type 3. Type 1 MNV is a blood vessel that originates from the capillary layer of the choroid and grows into the subretinal space of the RPE; type 2 MNV, formerly known as “classic CNV”, refers to the proliferation of neovascularization originating from the choroid that passes through the RPE and enters the subretinal space[50].
Scg3 has been identified as a disease-specific angiogenic factor in the retina, functioning independently of VEGF signaling. LeBlanc et al. suggested a potential role for Scg3 in CNV, showcasing its ability to stimulate endothelial cell growth and movement, thereby regulating angiogenesis and vascular permeability [51]. It is noteworthy that Scg3 expression levels were comparable in laser-induced CNV (LCNV) and healthy choroids. Through ligandomics, Ji et al. further supported the involvement of Scg3 in CNV by demonstrating that Scg3 binds to CNV vessels over 935 times more than healthy choroidal capillaries [52]. Moreover, Ji et al. conducted a ligand-binding assay for matrix-induced CNV (MCNV), revealing a 6.0-fold increase in Scg3 phage CNV binding compared to control chorionic vasculature [19]. Intriguingly, VEGF binding to CNV was similar to controls, suggesting that in the MCNV mouse model, the heightened binding of Scg3 to choroidal vasculature was specific compared to healthy choroidal capillaries within the same eye, with minimal induction triggered by the Scg3 ligand itself.
The MCNV model was induced in Scg3 knockout mice (Scg3-/-) and Scg3 wild-type mice (Scg3 + / +). The results demonstrated a significant reduction in CNV leakage intensity and area, as well as CNV 3D volume and largest lesion in Scg3-/- mice compared to Scg3 + / + mice. This suggests a disease-selective role for Scg3 in choroidal neovascularization [19]. These findings indicate that Scg3 functions as a disease-selective angiogenic factor not only in retinal pathology but also in choroidal neovascularization.
Anti-Scg3 inhibitorsCurrently, VEGF remains the primary target for anti-vascular therapy. However, inhibiting VEGF signaling in the retina may lead to unintended side effects, including arterial thromboembolism, stroke, and ocular inflammation [53]. An increasing body of evidence suggests that anti-Scg3 inhibitors might offer a safer and more effective alternative to anti-VEGF therapies. Numerous studies in recent years have explored anti-Scg3 inhibitors [17, 37, 38]. Scg3 promotes endothelial cell proliferation, migration, and tubule formation, yet its specific receptors have not yet been identified. The MEK/ERK signaling pathway emerges as crucial for Scg3-induced angiogenesis. MEK inhibitors effectively block Scg3-induced angiogenesis, while ERK inhibitors inhibit Scg3-induced endothelial cell proliferation and migration. Building upon the mechanism of action of these signaling pathways, studies have demonstrated that MEK and ERK inhibitors can significantly reduce neovascularization by impeding the transduction of signaling pathways [32].
Anti-Scg3 inhibition of retinal neovascularizationStudies have demonstrated that Scg3 activates the ERK/SRC signaling pathway, thereby modulating angiogenesis and vascular leakage [32]. Anti-Scg3 monoclonal antibody (anti-Scg3mAb) blocks its interaction with the cognate receptor, thus inhibiting Scg3-stimulated cell signaling. LeBlanc et al. investigated the impact of Scg3 inhibition on C57BL/6 J mice with diabetic retinopathy using a Scg3 monoclonal antibody [17]. In a mouse model of diabetic retinopathy, only pathological retinal neovascularization was inhibited, while retinal vascular leakage was reduced, and normal capillaries around the retina remained unchanged [42]. The ML49.3 monoclonal antibody recognizes human and mouse Scg3, and intravitreal injection of Scg3-neutralizing monoclonal antibody attenuates retinal vascular leakage in STZ-induced diabetic mice. Anti-Scg3 treatment was shown to attenuate diabetic retinal vascular leakage. Subsequent studies independently validated the therapeutic efficacy of Scg3 monoclonal antibody. In Ins2Akita mice with missense mutations in the insulin 2 gene spontaneously developing type 1 diabetes, mice developed retinal vascular leakage at 6 months of age. Intravitreal injection of anti-Scg3mAb in Ins2Akita mice significantly inhibited retinal leakage, confirming Scg3 as a target for DR therapy[54]. These findings suggest that anti-Scg3 monoclonal antibodies hold great potential as a novel therapy for the treatment of retinal neovascularization.
LeBlanc et al. developed ML49.3 and ML78.2mAb characterized as two non-competitive Scg3-neutralizing antibodies. Highly effective at alleviating OIR, another animal model of ROP, and inhibiting OIR-induced pathological retinal neovascularization [17, 36, 55]. Scg3 has been identified as a target for anti-angiogenic therapy. Intravitreal administration of ML78.2mAb confirmed a reduction in the intensity of OIR-induced RNV and neovascular clusters, facilitating the restoration of impaired vasculature to a normal state. Furthermore, to mitigate potential artifacts associated with intravitreal injection, intraperitoneal administration of ML78.2mAb significantly inhibited OIR-induced RNV, leading to decreased neovascular clusters and branch points[36]. Additionally, anti-Scg3hab therapy not only reduces pathological RNV and vascular leakage by selectively targeting the deep plexus of DR stress within the same diabetic retina but also facilitates blood reestablishment in the central avascular retina[43]. Improvements in arterial curvature and venous dilatation were observed, significantly reducing the severity of OIR as confirmed by fluorescein fundus angiography [37]. Using a mouse model, they found that the therapeutic range of anti-scg3hfab was at least tenfold wider than that of aflibercept [56]. In safety studies, anti-Scg3mab and anti-Scg3hab do not adversely affect retinal function. In contrast, aflibercept significantly inhibits normal retinal neovascularization. Not only does it not increase body weight in mice, it also has no adverse effect on the kidneys [36,37,38, 56]. Therefore, systemic safety was also evaluated, and the leakage of the Scg-3 inhibitor from the eye raised minimal safety concerns. However, the effectiveness and safety of this new therapy have not been fully explored in clinical trials.
Anti-Scg3 inhibition of choroidal neovascularizationThe research findings indicate that both anti-Scg3pAb and anti-Scg3mAb significantly reduced laser-induced choroidal neovascular leakage, CNV volume, lesion area, and vessel density [18, 20]. These results highlight the inhibitory effect of anti-Scg3 monoclonal antibody on choroidal neovascularization. In a recent study exploring anti-Scg3 gene therapy for the treatment of choroidal neovascularization in mice, it was observed that an adeno-associated virus-8 (AAV8) vector expressing anti-Scg3 Fab effectively ameliorated CNV with a comparable efficacy to that of AAV-aflibercept in a mouse model [57]. This study marks the first instance of testing an anti-angiogenic gene therapy. The therapeutic effects of anti-Scg3mab imply the potential induction of Scg3R in CNV, suggesting that anti-angiogenic drugs targeting disease-selective angiogenic receptors or their associated ligands may incur minimal side effects. Validation of the neutralizing activity of anti-Scg3hFab was further conducted with an in vitro functional assay, demonstrating its ability to block Scg3-induced human umbilical vein endothelial cell (HUVEC) migration and tubularization [19, 51]. Intravitreal injection of anti-Scg3hFab resulted in a significant reduction in the maximum area and volume of CNV lesions, while Scg3 deficiency nullified the effect of anti-Scg3hFab [51]. In a similar study, Ji et al. illustrated that even a small dose of anti-Scg3hFab could yield significant effects, as anti-scg3hfab at 0.5 µg/eye was more effective than the same dose of aflibercept in reducing the extent of CNV lesions. Notably, co-administration of anti-scg3hfab or aflibercept led to significant improvements in matrix-induced CNV (MCNV) in terms of CNV leakage and vessel staining, compared to administration of anti-scg3hfab or aflibercept alone [19]. Thus, Scg3 inhibitors offer a broader therapeutic window and lower drug rates, potentially enhancing their efficacy in treating retinal neovascularization. Nonetheless, further research is required to comprehensively establish their safety and toxicity profile.
Anti-Scg3 inhibition of corneal neovascularizationJin et al. conducted a study using a rabbit corneal alkali burn model to explore the effects of Scg3 on corneal neovascularization. Their research identified the presence of Scg3 expression in the corneal neovascularization of rabbits [
留言 (0)