
記住我





Mutations in SYNGAP1, a protein enriched at glutamatergic synapses, cause intellectual disability associated with epilepsy, autism spectrum disorder, and sensory dysfunctions. Several studies showed that Syngap1 regulates the time course of forebrain glutamatergic synapse maturation; however, the developmental role of Syngap1 in inhibitory GABAergic neurons is less clear. GABAergic neurons can be classified into different subtypes based on their morphology, connectivity, and physiological properties. Whether Syngap1 expression specifically in parvalbumin (PV)-expressing and somatostatin (SST)-expressing interneurons, which are derived from the medial ganglionic eminence (MGE), plays a role in the emergence of distinct brain functions remains largely unknown. We used genetic strategies to generate Syngap1 haploinsufficiency in (1) prenatal interneurons derived from the medial ganglionic eminence, (2) in postnatal PV cells, and (3) in prenatal SST interneurons. We further performed in vivo recordings and behavioral assays to test whether and how these different genetic manipulations alter brain function and behavior in mice of either sex. Mice with prenatal-onset Syngap1 haploinsufficiency restricted to Nkx2.1-expressing neurons show abnormal cortical oscillations and increased entrainment induced by 40 Hz auditory stimulation but lack stimulus-specific adaptation. This latter phenotype was reproduced in mice with Syngap1 haploinsufficiency restricted to PV, but not SST, interneurons. Prenatal-onset Syngap1 haploinsufficiency in Nkx2.1-expressing neurons led to impaired social behavior and inability to extinguish fear memories; however, neither postnatal PV- nor prenatal SST-specific mutant mice show these phenotypes. We speculate that Syngap1 haploinsufficiency in prenatal/perinatal PV interneurons may contribute to cortical activity and cognitive alterations associated with Syngap1 mutations.
Significance StatementMutations in the human gene cause a form of developmental epileptic encephalopathy associated with intellectual disability, autism, and sensory dysfunctions. Several studies have shown that in addition to playing a major role in the synaptic maturation and plasticity of forebrain excitatory neurons, Syngap1 affects GABAergic circuit function as well. Forebrain GABAergic neurons can be divided into different subtypes. Whether Syngap1 expression specifically in distinct interneuron populations and during specific developmental time windows plays a role in the emergence of distinct brain functions remains largely unknown. Here, we report that early, prenatal, or perinatal Syngap1 expression in developing GABAergic neurons derived from the medial ganglionic eminence promotes the development of auditory cortex function, social behavior, and ability to extinguish fear memories.
IntroductionSYNGAP1 codes for a Ras GTPase-activating protein that is highly abundant in the postsynaptic density of excitatory synapses (Komiyama et al., 2002; Kim et al., 2003; Vazquez et al., 2004). Work spanning two decades led to the discovery of multiple functions of Syngap1, including the regulation of plasticity-associated signaling pathways, structural roles in regulating postsynaptic density composition at cortical glutamatergic synapses (Komiyama et al., 2002; Kim et al., 2003; Vazquez et al., 2004; Clement et al., 2012; Ozkan et al., 2014; Araki et al., 2015, 2020; Zeng et al., 2016), and, more recently, nonsynaptic function in cortical neurogenesis (Birtele et al., 2023). In children, SYNGAP1 haploinsufficiency causes a developmental epileptic encephalopathy characterized by moderate to severe intellectual deficiency, generalized epilepsy (Hamdan et al., 2009; Rauch et al., 2012; Berryer et al., 2013; Carvill et al., 2013; von Stulpnagel et al., 2015; Mignot et al., 2016), and behavioral anomalies, including autism spectrum disorder and sensory processing impairment (Hamdan et al., 2011; Berryer et al., 2013; Michaelson et al., 2018; Carreno-Munoz et al., 2022; Lyons-Warren et al., 2022; Wright et al., 2022). Recent estimates suggest that SYNGAP1-ID may represent one of the most common monogenic forms of intellectual disability (Hamdan et al., 2009; Berryer et al., 2013, 2016; Parker et al., 2015), therefore making SYNGAP1 haploinsufficiency one of the most frequent causes of genetically defined childhood brain disorders. How Syngap1 regulates the development of different brain circuits and how its haploinsufficiency may contribute to specific phenotypes are still open questions and the focus of ongoing studies.
Syngap1 haploinsufficiency is associated with premature excitatory synaptic maturation in the first postnatal weeks of mouse cortical development, which could in turn affect activity-dependent postnatal refinement of cortical circuits and alter long-range circuit connectivity (Clement et al., 2012, 2013; Aceti et al., 2015). Furthermore, Syngap1 haploinsufficiency restricted to forebrain glutamatergic neurons is sufficient to recapitulate several of the phenotypes observed in Syngap1+/− mice, such as increased seizure threshold, hyperlocomotion, and deficits in working memory (Komiyama et al., 2002; Kim et al., 2003; Ozkan et al., 2014; Berryer et al., 2016; Creson et al., 2019; Nakajima et al., 2019). While Syngap1 expression appears to be enriched in cortical and hippocampal glutamatergic neurons, it is also expressed in GABAergic neurons (Zhang et al., 1999; Berryer et al., 2016; Su et al., 2019; Zhao and Kwon, 2023; Allen Brain Institute 10x Genomics human tissue). Consistent with this observation, mounting evidence suggests that Syngap1 affects the development and function of GABAergic inhibitory interneurons. For example, Syngap1 was shown to be essential for the proper migration of inhibitory neurons to dorsal cortices during development (Su et al., 2019). Furthermore, early postnatal Syngap1 deletion in sparse parvalbumin (PV)-expressing GABAergic cells in otherwise wild-type cortical organotypic cultures leads to a significant reduction of their perisomatic bouton density, suggesting a cell-autonomous role of Syngap1 in the maturation of PV interneuron synaptic innervation (Berryer et al., 2016). Furthermore, mice where Syngap1 haploinsufficiency is restricted to GABAergic interneurons, using the vesicular GABA transporter (Vgat) promoter to drive Cre expression, show impaired sensory learning in a whisker-guided tactile go/no-go task (Zhao and Kwon, 2023). Finally, we previously observed that mice where Syngap1 haploinsufficiency is restricted to neurons derived from the medial ganglionic eminence (MGE), including PV- and SST-expressing interneurons, show behavioral deficits in the social domain (Berryer et al., 2016). Cortical GABAergic interneurons can be divided into different populations based on their gene expression profile, connectivity, and physiological properties, and this diversity likely contributes to different functional roles in cortical circuits (Kepecs and Fishell, 2014; Tremblay et al., 2016). Whether Syngap1 differently affects different interneuron types is unknown. Furthermore, whether Syngap1 function in GABAergic interneurons is limited to early development or extends to later stages is unclear. Here, we address these questions by using different conditional transgenic mouse strategies, in vivo recordings, and behavioral assays.
Materials and MethodsMiceAll procedures and experiments were done in accordance with the Comité Institutionnel de Bonnes Pratiques Animales en Recherche of the CHU Sainte-Justine Research Center in line with the principles published in the Canadian Council on Animal Care. Syngap1lox mice were purchased from the Jackson Laboratory (#029303). All animals were maintained under a light/dark cycle (12 h light/dark) in a temperature- and humidity-controlled room. Food and water were available ad libitum. Syngap1lox mice were crossed either with the Nkx2.1-Cre mice (Jackson Laboratory #008661), PV-Cre mice (Jackson Laboratory #008069), or SST-Cre (Jackson Laboratory #018973) mice. To quantify SST and PV interneuron density, we crossed these mice with mice carrying the RCE allele (Jackson Laboratory #032037). In the RCE allele, a STOP cassette flanked by the loxP sites was inserted upstream of the coding sequence of EGFP, thus allowing EGFP expression in Cre-expressing cells. The resulting genotypes were as follows: Tg(Nkx2.1-Cre);RCElox/lox;Syngap1+/+ (Nkx2.1-Cre control) and Tg(Nkx2.1-Cre);RCElox/lox;Syngap1lox/+ (Nkx2.1-Cre cHET). Tg(Nkx2.1-Cre);RCElox/lox;Syngap1lox/+ mice were further crossed together to obtain Tg(Nkx2.1-Cre);RCElox/lox;Syngap1lox/lox mice (Nkx2.1-Cre cKO) along with control and Nkx2.1-Cre cHET littermates. We also generated the following genotypes: Tg(PV-Cre);RCElox/lox;Syngap1+/+ (PV-Cre control), Tg(PV-Cre);RCElox/lox;Syngap1lox/+ (PV-Cre cHET), Tg(SST-Cre);RCElox/lox;Syngap1+/+ (SST-Cre control), and Tg(SST-Cre);RCElox/lox;Syngap1lox/+ (SST-Cre cHET). Mice of both sexes were used for all experiments except for fear conditioning, which was performed using males only.
GenotypingDNA was extracted from mouse tails using the AccuStart II Mouse Genotyping kit (#95135-500). Polymerase chain reaction (PCR) was done to detect the presence of Cre, RCE, Syngap1lox, and wild-type alleles. To detect the Syngap1lox alleles, two separate primers were used (F1 5′-GGGCTGTAAAACCCAACAAG-3′ and R1 5′-GCAGCTTTTTCTCAGGGAAC-3′) to detect a band size at 420 bp for the floxed allele and 372 bp for the WT allele. To detect the Nkx2.1-Cre transgene, three separate primers were used (F1 5′-AAGGCGGACTCGGTCCACTCCG-3′, F2 5′-TCCTCCAGGGGACTCAAGATG-3′, and R1 5′-TCGGATCCGCCGCATAACCAG-3′) to detect a band size at 220 bp for wild-type and 550 bp for the transgene. For PV-Cre transgene, three separate primers were used (F1 5′-CAGCCTCTGTTCCACATACACTCC-3′, F2 5′-GCTCAGAGCCTCCATTCCCT-3′, and R1 5′-TCACTCGAGAGTACCAAGCAGGCAGGAGATATC-3′) to detect a band size at 526 bp for wild-type and 400 bp for the transgenic allele. To detect the SST-Cre transgene, three separate primers were used (F1 5′-TCTGAAAGACTTGCGTTTGG-3′, F2 5′-TGGTTTGTCCAAACTCATCAA-3′, and R1 5′-GGGCCAGGAGTTAAGGAAGA-3′) to detect a band size at 465 bp for wild-type and 200 bp for the mutant allele. The RCE allele was detected using three separate primers (RCE-Rosa1 F1 5′-CCCAAAGTCGCTCTGAGTTGTTATC-3′, RCE-Rosa2 F2 5′-GAAGGAGCGGGAGAAATGGATATG-3′, and RCE-Cag3 R1 5′-CCAGGCGGGCCATTTACCGTAAG-3′) to detect band sizes at 550 and 350 bp for wild-type and mutant allele, respectively.
Immunofluorescence analysisP60 mice were anesthetized with [ketamine (100 mg/kg) plus xylazine (10 mg/kg) plus acepromazine (10 mg/kg)] and perfused transcardially with 0.9% saline followed by 4% paraformaldehyde (PFA) in phosphate buffer (0.1 M PB, pH 7.2–7.4). Brains were dissected out and postfixed in 4% PFA overnight at 4°C. They were subsequently transferred to 30% sucrose (prepared in PBS, pH 7.2) at 4°C for 48 h. Brains were then embedded in molds filled with OCT Tissue Tek and frozen in a bath of 2-methylbutane placed on a bed of dry ice and ethanol. Coronal sections were cut at 40 μm with a cryostat (Leica CM3050 S) and collected as floating sections in 1× PBS. Brain sections were blocked in 10% normal goat serum (NGS) with 1% Triton X-100 in PBS for 2 h at room temperature (RT) followed by incubation at 4°C for 48 h with the following primary antibodies diluted in 5% NGS and 0.1% Triton X-100 in PBS: rabbit anti-PV (1:5,000, Swant, catalog #PV27), rat anti-SST (1:500, Millipore, catalog #MAB354), mouse anti-NeuN (1:500, Millipore, catalog #MAB377), and chicken anti-GFP (1:500, Abcam, catalog #13970). Sections were then washed in PBS plus 0.1% Triton X-100 (3× 10 min each) and incubated for 2 h at RT with the following secondary antibodies diluted in 5% NGS and 0.1% Triton X-100 in PBS: Alexa 488-conjugated goat anti-chicken (1:1,000, Abcam, catalog #ab150169), Alexa 555-conjugated goat anti-rabbit (1:500, Life Technologies, catalog #A21430), Alexa 647-conjugated goat anti-mouse (1:500, Cell Signaling Technology, catalog #4410), Alexa 555-conjugated goat anti-rat (1:500, Life Technologies, catalog #A21434). Sections were rinsed in PBS (3× 10 min each) and mounted with Vectashield mounting medium (VectorLabs). Immunostained sections were imaged using a Leica SP8-DLS confocal microscope, with a 20× magnification (NA 0.75) at 1,024 × 1,024, zoom at 1, z-step of 1.5 μm, and stack size of ∼20 μm. Images were acquired from at least three coronal sections for each animal. Acquisition and analysis of images were done by an experimenter blind to the genotype. For neuron density quantification, 3–4 ROIs were drawn in each of the layers, and the number of cells was quantified using the cell counting option in Neurolucida (Microbrightfield Bioscience).
Fluorescence multiplex RNAscopeMice were cervically dislocated, and brains were dissected out at two ages, i.e., P20 and P60. Brains were washed with PBS and fast-frozen for ∼30–60 s in a cold bath of 2-methylbutane on a bed of dry ice and ethanol and preserved at −80°C. Twenty-micrometer coronal sections were cut with a cryostat (Leica Microsystems) and serially mounted (3–4 sections per slide) on Superfrost Plus Gold glass slides (Thermo Fisher Scientific catalog #12-550-15), which were then stored at −80°C until further use. RNAScope Multiplex Fluorescent Reagent Kit v2 for the reagents was purchased from Advanced Cell Diagnostics (catalog #320851) along with the following probes: Mm-Syngap1 - Mus musculus synaptic Ras GTPase-activating protein 1 homolog (rat) (Syngap1) for Syngap1 mRNA (catalog #417381), Mm-Pvalb-C2 - Mus musculus parvalbumin (Pvalb) mRNA for Pvalb mRNA (catalog #421931-C2), Mm-Sst-C2 - Mus musculus somatostatin (Sst) for Sst mRNA (catalog #404631-C2), positive probe (catalog #320881), and negative probe (catalog #320871). The entire procedure for tissue pretreatment, hybridization, amplification, and detection was performed as per the RNAscope Multiplex Fluorescent Assay (Advanced Cell Diagnostics) manual for fresh frozen tissue. For pretreatment, slides were removed from −80°C and immediately postfixed in 4% PFA at 4°C for 15 min followed by dehydration in 50, 70, and 2× 100% ethanol for 5 min, respectively, at room temperature (RT). The slides were air-dried, and a hydrophobic barrier was created around the section with an ImmEdge Pen (H-4000). Slides were treated with protease IV for 15 and 30 min, for P20 and P60 brain sections respectively, at RT, and then washed twice in PBS. All the probes were prewarmed at 40°C in a water bath for 10 min and cooled at RT. For hybridization, the probes were incubated at 40°C for 2 h. Amplification steps were carried out as per the protocol. For detection of the probes, Amp4B was used (C1 probes-Atto 550 and C2 probes-Alexa-488). Sections were stained with DAPI (320858) for 30 s and mounted with ProLong Diamond Antifade (Thermo Fisher Scientific P36961). Images of the primary cortical auditory cortex were taken with an SP8-STED confocal microscope using a 63× magnification (NA 1.4) at 2,048 × 2,048, zoom of 2, and z-step of 0.3 μm with gating between 0.3 and 6.0 ms. At least three brain slices/mouse were imaged. Images were deconvolved using Huygens HyVolution developed for Leica Systems. Quantification of mRNA was performed using Fiji (ImageJ), as described by Lavertu-Jolin et al. (2023). Putative cell somata were manually selected by using DAPI staining and either Pvalb or Sst puncta distribution. Cells were duplicated, separated into channels, and filtered using 3D Gaussian blur. Dots were quantified by using the 3D object counter in ImageJ Fiji, after setting an appropriate threshold value for each of the probes. Brain slices from P20 and P60 mice were processed at the same time. The results were reported as the mean number of dots per cell.
Behavioral analysisMice of either sex, unless differently specified, were housed under a 12 h light/dark cycle in a temperature- and humidity-controlled room. Food and water were available ad libitum. Mice were habituated to the operator prior to all behavior experiments. All behavior experiments were performed with the light intensity range of 60–100 lux. A camera was mounted above the arena for video recording during the behavioral task and controlled by SMART video tracking software (Panlab v3.0, Harvard Apparatus) or Freeze Frame software IMAQ 3 (Version 3.0.1.0). The sequence of tested mice was randomized by the genotype. Mice were tested between P60 and P90. Each behavioral apparatus was cleaned with 70% ethanol between each trial. Experimenters were blind to genotypes during both testing and data analysis. All behavioral experiments were done using at least 3–4 cohorts from different mouse litters. If not differently specified, we use both male and female mice for these studies.
Open fieldEach mouse was gently placed in a corner of the open field apparatus (box dimension, 45 × 45 cm) and allowed to explore for 10 min. No mice were excluded from the analysis.
Elevated plus mazeThe apparatus consists of two open arms without walls facing across each other and perpendicular to two closed arms with walls joining at the central platform. Each mouse was placed at the center of the junction of two open and closed arms and allowed to explore for 5 min. No mice were excluded from the analysis.
Novel object recognitionEach mouse was habituated to the open field box for a duration of 5 min. For the familiar phase, 1 h after habituation, two familiar objects (two similar cylindrical objects: white in color) were placed in the box at one-third the distance from one end of the box. Each mouse was placed at one end of the box facing the wall away from the objects and allowed to explore for 10 min. Only mice that explored the objects for at least 10 s were considered for further analysis. Based on this criterion, we excluded two Nkx2.1-Cre cHET and two Nkx2.1-Cre Ctrl mice. The discrimination index (DI) was calculated as (Time spent with Object 1 − Time spent with Object 2) / (Total time spent with Object 1 + Object 2). Two hours later, one of the familiar objects was replaced with a novel object (cone-shaped object; white in color). Each mouse was allowed to explore for 10 min, and DI was calculated as (Time spent with novel object − Time spent with familiar object) / (Total time spent with novel object + familiar object). The placement of the familiar and the novel objects was randomized for each mouse.
Three-chamber testEach mouse was placed in the center of a three-chamber rectangular box (dimension, 61 × 42.5 cm) and allowed to explore for 10 min (habituation). Next, a wire cage with an unfamiliar conspecific mouse of the same size, age, and sex (Stranger 1) was placed on one side of the chamber, while an empty wire cage was placed on the other side of the chamber. The test mouse was placed in the center of the chamber and allowed to explore for 10 min (social approach). Finally, a novel unfamiliar mouse (Stranger 2) of the same size, age, and sex was placed in the previously unoccupied empty cage, and the test mouse was allowed to explore for 10 min (social novelty). Strangers 1 and 2 were taken from different home cages and have never been in close physical contact with the test mouse or with each other. No mice were excluded from the analysis.
Contextual fear conditioningMale mice were habituated on Day 1 and allowed to freely explore for 3 min in the test context, which consisted of white walls and a grid floor for habituation. On Day 2, mice were fear conditioned using three random unconditioned stimuli (US; 2 s footshock, 0.5 mA), delivered over a period of 5 min. Mice were tested in the same context both 24 h and 30 d later. An experimenter blind to the genotype scored freezing behavior defined by the time the mouse remained in complete immobility with the exception of respiratory movements, during the 5 min period spent in the context. No mice were excluded from the analysis.
Cued fear conditioning and fear extinctionFear acquisition and extinction experiments were performed as described by Lavertu-Jolin et al. (2023), using two different contexts (Context A and Context B, respectively). Context A consisted of white walls and a grid floor and was cleaned with 70% ethanol before and after each session. Context B consisted of walls with black and white stripes and a white plexiglass floor and was cleaned with 1% acetic acid before and after each session. On Day 1, mice were allowed to freely explore for 3 min in Context A (habituation). On Day 2, mice were conditioned with five random pairings of the conditioned stimulus (CS; duration, 5 s; white noise, 80 dB) with coterminating US (2 s footshock, 0.6 mA; intertrial interval, 30–60 s). On Days 3 and 4, mice were subjected to early and late extinction training in Context B with 12 presentations of CS at 30 s intervals on each day. Fear retrieval and renewal were tested 7 d later in Contexts B and A, respectively, using four CS presentations at an interval of 30 s. Mouse behavior was video-recorded with Freeze Frame software. An experimenter blind to the genotype scored freezing behavior (defined by complete immobility with the exception of respiratory movements) by measuring time spent freezing for 30 s following CS presentation for extinction and during 1 min following CS presentation in retrieval and renewal. The results presented are the mean of time spent freezing after the presentation of the first two CS on the first day of extinction (Day 2; CS1–2, early extinction), the last two CS on the second day of extinction (Day 3; CS 11–12, late extinction), the first two CS during the retrieval test in the extinction Context B (Day 10; CS 1–2, retrieval), and the first two CS during the renewal test in the fear acquisition Context A (Day 10; CS 1–2, renewal). No mice were excluded from the analysis.
In vivo electrophysiologySurgeryFor EEG recordings in anesthetized mice, anesthesia was induced by an intraperitoneal injection of urethane using an initial large dose (1.5 g/kg, i.p.) followed by supplementary small doses (0.1–0.15 g/kg) during the recording, if necessary, as indicated by the presence of a withdrawal response of the limb to pinch. The animal was transferred to the stereotaxic apparatus for both surgery and subsequent recording. A craniotomy was performed on the right primary auditory cortex of the brain at the following coordinates: −2.5 mm posterior to bregma, 3.9 mm lateral to the midline, and 0.8 mm deep. A second craniotomy was performed in the contralateral prefrontal lobe where the reference and ground reference wire were placed. Custom-made electrodes were made with a cluster of five 50-µm-diameter insulated tungsten wires. Wire tips were 100 µm apart to reach different depths. Body temperature was maintained at 36–37°C by a thermostatically controlled heating pad.
For EEG recordings in awake mice, electrodes were implanted under deep anesthesia, induced by an intraperitoneal injection of ketamine/xylazine/acepromazine cocktail (80 mg/kg ketamine, 15 mg/kg xylazine, and 2 mg/kg acepromazine). A craniotomy was performed on the right side of the primary auditory cortex as described for recording in anesthetized mice. Custom-made clusters of four to six 50-μm-diameter insulated tungsten wires were implanted. Wire tips (recording sites) were 50–150 μm apart. A ground-and-reference wire was also gently introduced in the contralateral frontal lobe, and a 2 cm carbon fiber bar was placed on the back of the skull. The whole structure was then daubed with dental acrylic to encase the electrode–microdrive assembly and anchor it to the skull. During the whole surgery, body temperature was maintained at 36–37°C by a thermostatically controlled heating pad. For a week after surgery, animals were treated with Metacam, and the skin around the implant was disinfected daily for 3 d following the surgery. Experimenters were blind during surgery, data acquisition, and analysis.
EEG recordingEEG recordings were performed using an Open Ephys GUI platform (https://open-ephys.org/) at a sampling rate of 20 kHz. Data were acquired with a custom-made headstage using Intan Technologies’ RHD Electrophysiology Amplifier Chip. EEG data and video were recorded simultaneously. Custom-designed stimulus trigger generator devices sent triggers directly to the recording system assuring the time precision of the stimulus presentation.
Sensory stimulations and analysisThese experiments were performed as described by Carreno-Munoz et al. (2022). All stimulations were presented at an intensity of 70 dB. For pure tone stimulation, we used 100-ms-long 5 and 10 kHz pure tones presented with a random interstimulus interval of 2–3 s. Each pure tone was presented 60 times in random order. The full protocol lasted 5 min. For the habituation/oddball paradigm, we used sequences of 5–9 repetitive (standard, at 5 kHz) sounds followed by a deviant sound (at 10 kHz). Sequences were presented in a pre-established pseudorandom order. Sounds lasted 70 ms and were presented at 1 s interstimulus interval. The full protocol lasted 10 min. For auditory steady-state stimulation, we used 10 and 40 Hz click trains (1 s duration and 2.5 s intertrial interval), presented at alternating frequencies. Each click was a sound at 5 kHz, lasting 5 ms. This protocol lasted 5 min.
EEG preprocessingThe mouse EEG signals were downsampled to 2,000 Hz and filtered with a passband filter (0.5–150 Hz) and a notch filter (59.5–60.5 Hz) to remove residual 60 Hz powerline noise contamination. Data were then segmented into periods of different lengths, depending on the stimulation protocol: 1,400 ms (500 ms pre- and 900 ms poststimulus onset) for the auditory oddball protocol; 2,500 ms (1,000 ms pre- and 1,500 ms poststimulus onset) for the pure tone stimulation; and 4,000 ms (1,000 ms pre- and 3,000 ms poststimulus onset) for auditory steady state. Trials containing per-sample segments with an intrachannel average >4 times the total trial standard deviation were tagged, visually inspected, and removed. Only sessions containing >50 clean trials were kept for further analyses. Signal analysis and quantification was performed using custom MATLAB (MathWorks) code, available upon request.
Auditory-evoked potentialsThe EEG signal was low-pass filtered to 150 Hz, baseline corrected to the mean voltage of the 150 ms prior to stimulus onset, and averaged over the trial. Event-related potential components baseline-to-peak N1 and P1 were analyzed from the AEP. The N1 amplitude was automatically detected by subtracting the minimum voltage (negative peak) within a 10–80 ms time window after stimulus onset to the averaged baseline value. The P1 amplitude was extracted by subtracting the maximum voltage (positive peak) within an 80–150 ms time window after stimulus onset to the averaged baseline value. Analyses were carried out using FieldTrip toolbox v. 202009.
Habituation and mismatch negativityTo quantify habituation in mice, we calculated the ratio between the amplitudes of the first standard sound (S1) and the second (S2). These ratios were then converted to a logarithmic base 10 scale. Mismatch negativity (MMN) was calculated by subtracting the EEG trace of the standard sound from the EEG trace of the deviant sound. The pattern obtained was then used to detect N1 and P1 amplitudes in each subject.
Intertrial coherence. Intertrial coherence allows assessment of the strength of phase coherence across trials in temporal and spectral domains. The intertrial coherence computation uses only the phase of the complex values given by Morlet's wavelet transform. Intertrial coherence measures phase coupling across trials at all latencies and frequencies and is defined as follows:ITCt=1N∑n=1Nexp(jΘ(f,t,n)), where jΘ(f,t,n) represents the phase for a given frequency (f), time point (t), and trial (n). The obtained values are always defined between 0 and 1. Phase-locking values close to 1 indicate strong intertrial phase-locking, thus representing evoked activity while scores closer to 0 indicate a high intertrial phase variability.
Experimental design and statistical analysisFor each experiment, littermates of both genotypes, from at least three different litters, were analyzed. Data were first tested for normal distribution with D’Agostino and Pearson's test, Anderson–Darling test or Shapiro–Wilk test, and Kolmogorov–Smirnov test. Data involving only two experimental groups were analyzed using either (1) t test with Welch's correction for normally distributed data or (2) Mann–Whitney test for non-normally distributed data. Data with >2 experimental groups were analyzed using one-way ANOVA with post hoc Dunn's multiple-comparisons test for normally distributed data or Kruskal–Wallis test for non-normally distributed data. For the three-chamber test (social behavior), sniffing times were compared using two-way ANOVA with post hoc Holm–Sidak's multiple-comparisons test.
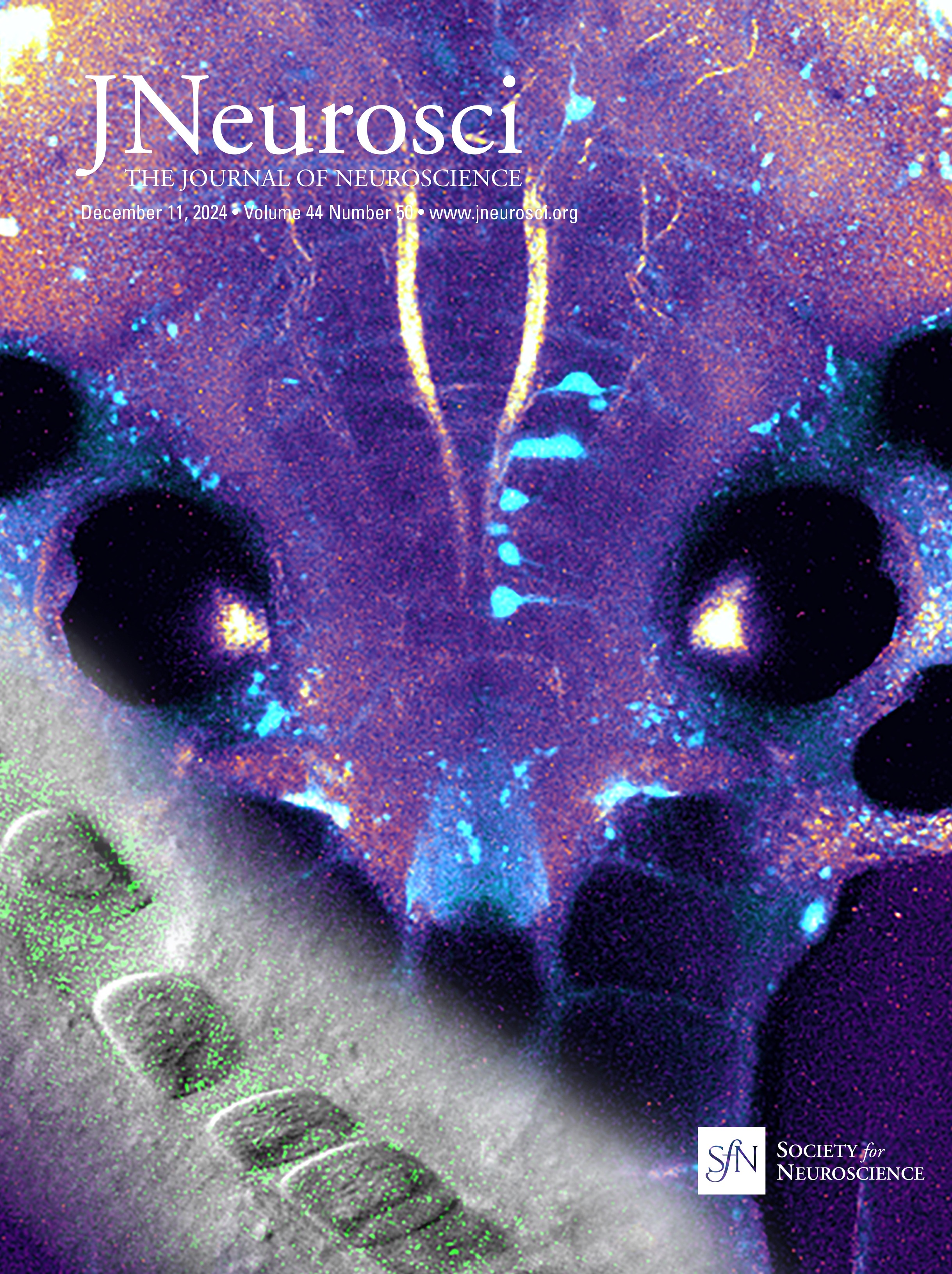
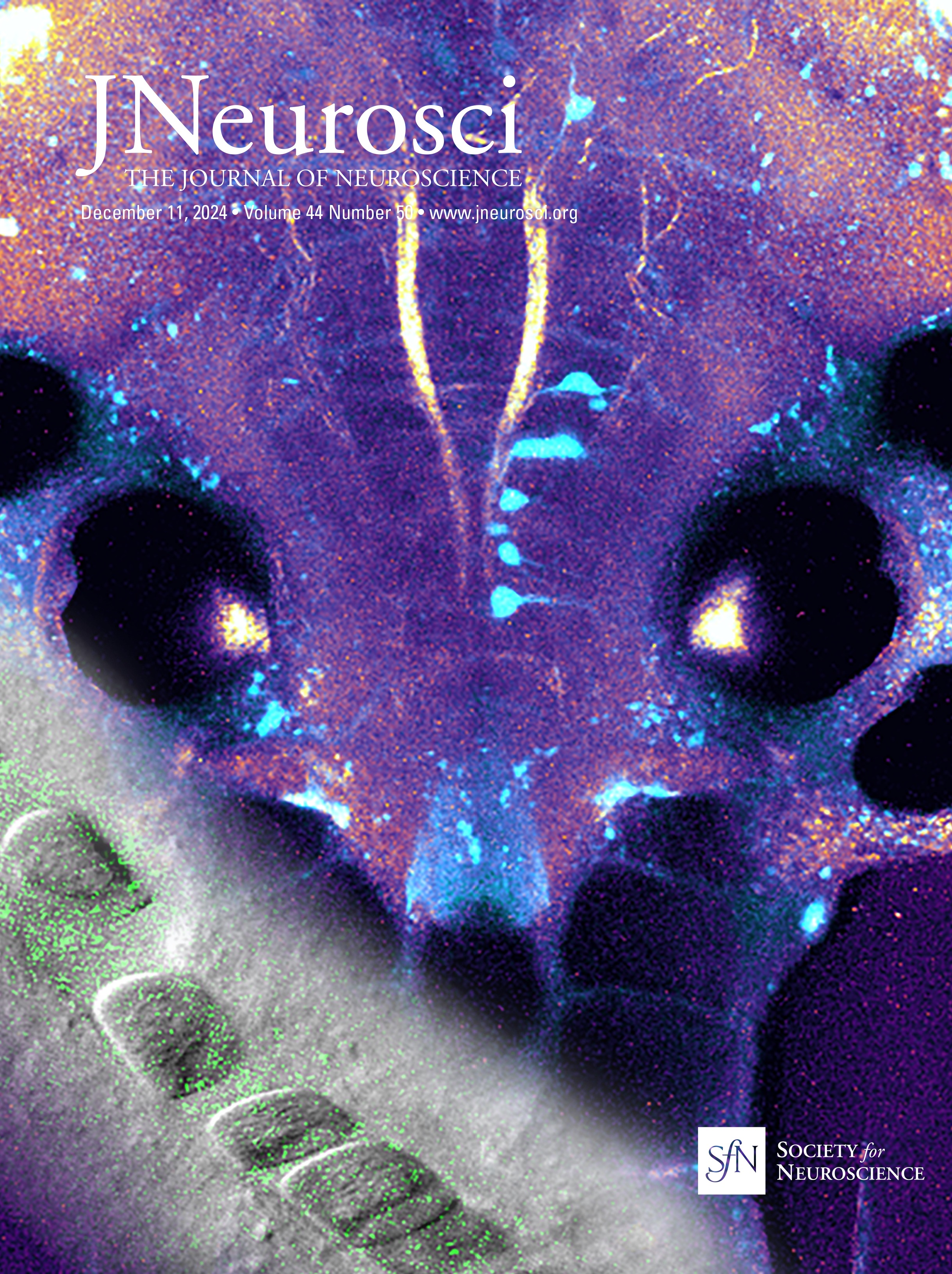
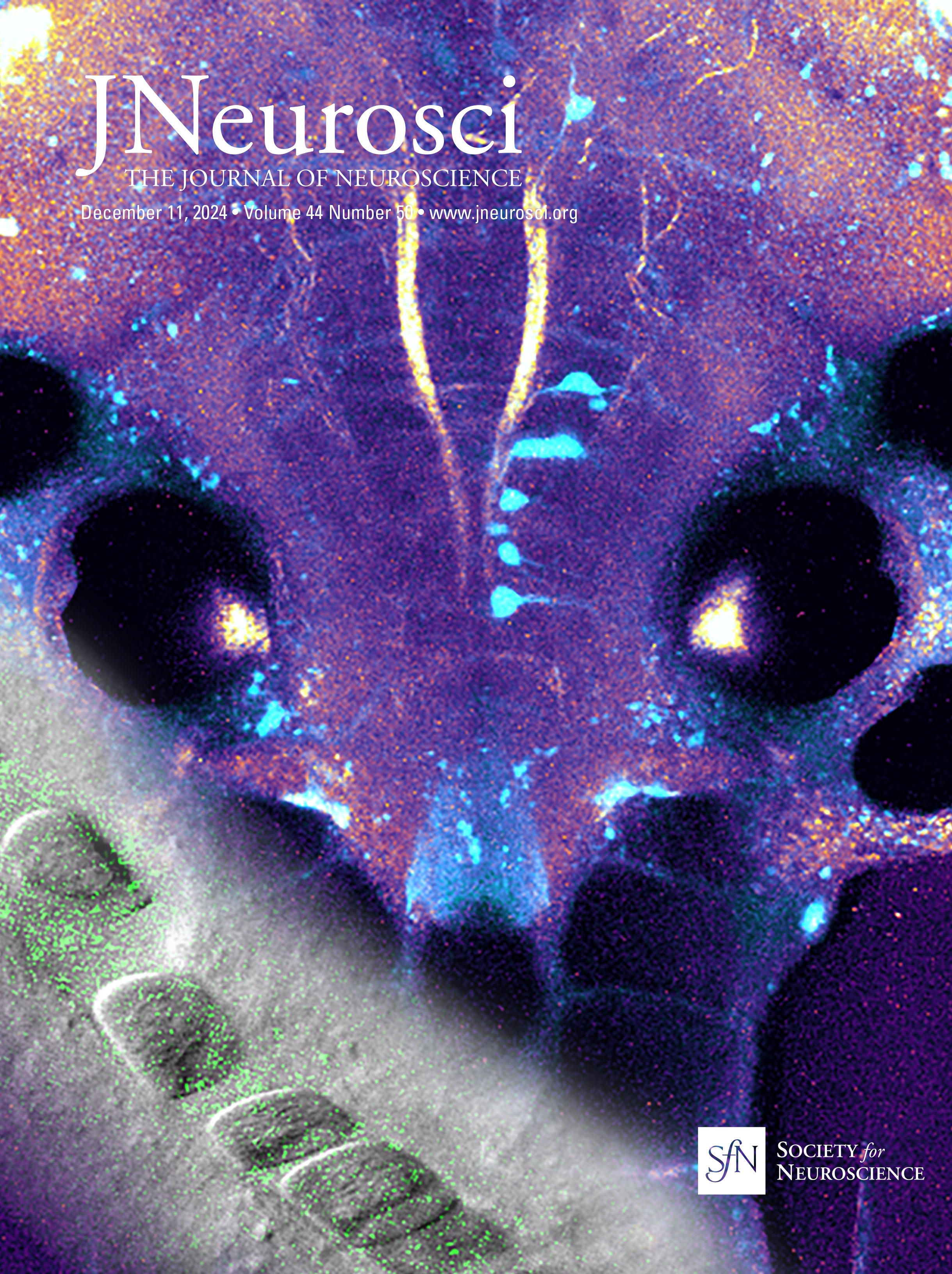
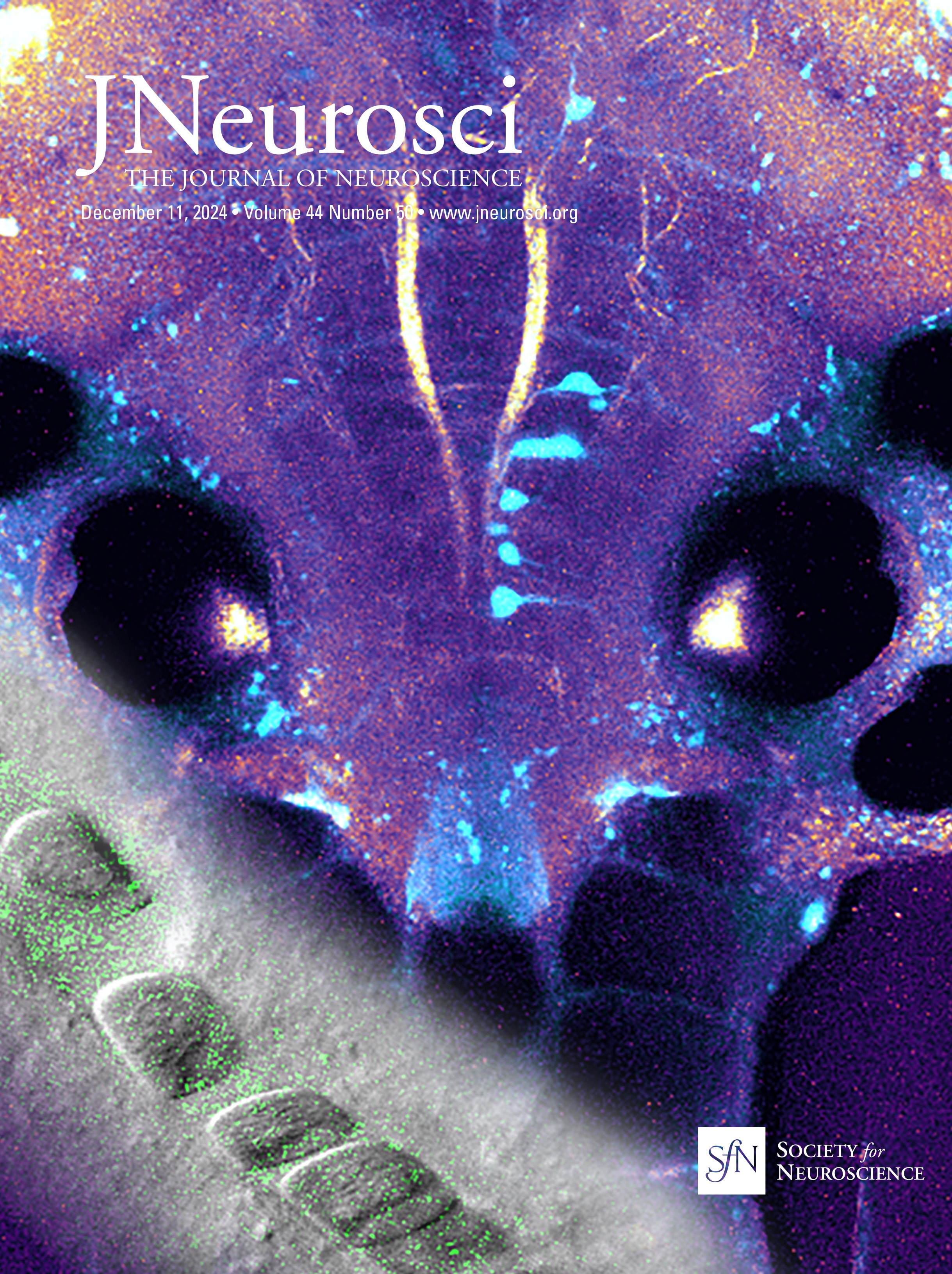
ResultsSyngap1 mRNA is expressed at higher levels in preadolescent than in adult PV+ and SST+ cortical interneuronsSyngap1 expression has been detected in a subset of GABAergic neurons in dissociated hippocampal and cortical neuronal cultures (Zhang et al., 1999; Moon et al., 2008; Berryer et al., 2016). Previous analysis of Syngap1 expression in cortical interneuron subtypes using transcriptome datasets collected by the Allen Brain Institute suggests that Syngap1 is detected in GABAergic interneurons and excitatory cells in the adult cortex (Zhao and Kwon, 2023). We first sought to confirm Syngap1 expression in cortical PV and SST neurons and then investigated whether its expression may be different in the developing versus the adult cortex. Since Syngap1 immunostaining in mouse brain slices gives a diffuse signal with poor spatial resolution, we turned to RNAscope Multiplex Fluorescent in situ hybridization (Advanced Cell Diagnostics), which allows single-mRNA molecule detection. We simultaneously labeled the mRNA coding for SYNGAP1 (gene name, Syngap1) and either PV (gene name, Pvalb) or Somatostatin (gene name, Sst) in the cortex of preadolescent (P20) and young adult (P60) mice (Fig. 1A,B). We chose the two developmental points since cortical GABAergic connectivity undergoes a prolonged postnatal maturation phase, accelerating after the first postnatal week and reaching a plateau by the first postnatal month (Micheva and Beaulieu, 1996; Chattopadhyaya et al., 2004; Pangratz-Fuehrer and Hestrin, 2011; Favuzzi et al., 2019). We found that Pvalb- and Sst-positive interneurons expressed Syngap1 at both ages. However, Syngap1 mRNA particle numbers were significantly higher at P20 than at P60 in both GABAergic cell populations (Fig. 1D,F). This was not due to age-dependent changes in mRNA detection, since both Pvalb and Sst mRNA particle numbers were not significantly different at P20 versus P60 (Fig. 1C,E). To investigate whether the decrease in Syngap1 expression was specific to GABAergic cells or a widespread phenomenon, we quantified Syngap1 mRNA in Pvalb-negative neurons, which, most likely, represented pyramidal neurons, and found a similar age-dependent decrease (Fig. 1G). Of note, Syngap1 mRNA particle numbers were significantly higher in Pvalb- than in Sst-positive interneurons at P20, but the difference did not reach significance at P60 (P20, Welch's t test, p = 0.0227; P60, Welch's t test, p = 0.0597; number of mice, n = 5 for both ages). When comparing Syngap1 mRNA particle numbers Pvalb-positive versus Pvalb-negative (most likely excitatory neurons), we found that the latter showed a trend toward higher Syngap1 expression (P20, Welch's t test, p = 0.0648; P60, Welch's t test, p = 0.0544; number of mice, n = 5 for both ages). These data suggest that PV neurons express higher levels of Syngap1 mRNA; however, whether different mRNA particle number translates in different protein expression levels remains to be investigated.
Syngap1 haploinsufficiency in MGE-derived Nkx2.1-expressing interneurons does not alter interneuronal density or final positioning in the adult cortexTo investigate the role of Syngap1 in the early development of PV and SST interneurons, we used Nkx2.1Cre;RCE;Syngap1lox/+ mice to remove either one or both Syngap1 alleles starting at around embryonic day (E) 10.5. Nkx2.1, a transcription factor expressed in the medial ganglionic eminence (MGE), is essential for the specification of cortical PV and SST interneurons (Q. Xu et al., 2004, 2008), while RCE is a reporter line expressing eGFP in a Cre-dependent manner. In this mouse line, we confirmed that the proportion of PV and SST neurons expressing GFP varied depending on the cortical layer and brain region. In particular, the proportion of PV cells expressing GFP was 66 ± 2% and 84 ± 2% in sensory cortex L23 and L56, respectively, and 88 ± 3% in the prefrontal cortex of adult mice (N = 6 mice). The proportion of SST cells expressing GFP was 57 ± 4% and 72 ± 9% in sensory cortex L23 and L56, respectively, and 73 ± 5% in the prefrontal cortex of adult mice (N = 3 mice). Not all cortical SST and PV interneurons express Cre most likely because the Nkx2.1Cre transgenic mouse was generated not by knock-in of Cre downstream the endogenous Nkx2.1 promoter but by genomic integration of a modified bacterial artificial chromosome, in which the second exon of Nkx2.1 was replaced by Cre (Q. Xu et al., 2008).
 Figure 1.
Figure 1. Syngap1 transcripts are significantly reduced in PV, SST, and non-PV cells in adult compared with preadolescent mice. A, B, Representative RNAscope images of layer 5 auditory cortex from P20 and P60 mice probed with Pvalb (cyan) and Syngap1 (magenta; A) or Sst (cyan) and Syngap1 (magenta; B). Cell nuclei are labeled with DAPI (gray). Pvalb-positive (A) and Sst-positive (B) cell somata are indicated by yellow arrowheads, while Pvalb-negative (A) or Sst-negative somata (B) are indicated by white arrows. Scale bar, 5 µm. C, Pvalb transcript number per cell at P20 and P60 (Welch's t test, p = 0.1204). D, Syngap1 transcripts in Pvalb+ cells at P20 and P60 (Welch's t test, **p = 0.0064). E, Sst transcript number per cell at P20 and P60 (Mann–Whitney test, p = 0.5476). F, Syngap1 transcripts in Sst+ cells at P20 and P60 (Welch's t test, **p = 0.0010). G, Syngap1 transcripts in Pvalb− cells at P20 and P60 (Welch's t test, **p = 0.0018). Number of mice, n = 5 for both ages. Bar graphs represent mean ± SEM. p > 0.05, not significant (ns), *p < 0.05, **p < 0.01, ***p < 0.001.
Syngap1 has been shown to be involved in the migration of inhibitory neurons during development (Su et al., 2019); therefore, conditional prenatal deletion of Syngap1 may affect their final numbers in the adult cortex by altering their final positioning. Here, we used the Syngap1lox transgenic line reported previously (Clement et al., 2012; Ozkan et al., 2014) wherein the loxP sites flank Exons 6–7 (hereafter indicated simply as Syngap1lox). Neuron-type–specific Cre recombinase expression in this mouse line led to the efficient removal of Syngap1 protein in both glutamatergic and GABAergic cortical cells (Ozkan et al., 2014). Neither conditional heterozygous (Nkx2.1Cre;RCE;Syngap1lox/+ referred to hereafter as cHET) nor conditional homozygous (Nkx2.1Cre; RCE;Syngap1lox/lox referred to hereafter as cKO) mice showed significant difference in PV (Fig. 2A,B1–D1,B2–D2) and SST (Fig. 2E,F1–H1,F2–H2) neuron density compared with wild-type littermates (Nkx2.1Cre; RCE;Syngap1+/+ referred to hereafter as control) in sensory (Fig. 2B1–C2,F1–G2) and associative cortices (Fig. 2D1,D2,H1,H2). Therefore, Syngap1 deletion in Nkx2.1-expressing GABAergic progenitors does not appear to affect the survival and final positioning of adult cortical PV and SST interneurons.
 Figure 2.
Figure 2. Density and distribution of PV- and SST-positive cells are not affected by haploinsufficiency or deletion of Syngap1 in MGE-derived interneurons. A, Representative image of cortical coronal sections from control and MGE-derived conditional mutant mice (heterozygous, Nkx2.1-Cre cHET; homozygous, Nkx2.1 Cre cKO) immunostained for PV (magenta), GFP (cyan), and NeuN (blue). Scale bar, 50 µm. B1–D1, Percentage of PV+ GFP+ over NeuN+ cells in sensory cortex layer 2/3 (B1) and layer 5/6 (C1) and in the prefrontal cortex (D1). B1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.3862 for Nkx2.1Cre-cHET and p = 0.9450 for Nkx2.1Cre-cKO versus Ctrl. C1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.6095 for Nkx2.1Cre-cHET and p = 0.8389 for Nkx2.1Cre-cKO versus Ctrl. D1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.1456 for Nkx2.1Cre-cHET and p = 0.0857 for Nkx2.1Cre-cKO versus Ctrl. B2–D2, Density of PV+ GFP+ per square micrometer in sensory cortex layer 2/3 (B2) and layer 5/6 (C2) and in the prefrontal cortex (D2). B2, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.9546 for Nkx2.1Cre-cHET and p = 0.8474 for Nkx2.1Cre-cKO versus Ctrl. C2, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.5588 for Nkx2.1Cre-cHET and p = 0.7473 for Nkx2.1Cre-cKO versus Ctrl. D2, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.3033 for Nkx2.1Cre-cHET and p = 0.3698 for Nkx2.1Cre-cKO versus Ctrl. Number of mice, n = 6 Ctrl; n = 4 Nkx2.1Cre-cHET and n = 4 Nkx2.1Cre-cKO. E, Representative image of cortical coronal sections from control and MGE-derived conditional mutants of adult mice immunostained for SST (magenta), GFP (cyan), and NeuN (blue). Scale bar, 50 µm. F1–H1, Percentage of SST+ GFP+ normalized over NeuN+ cells in sensory cortex layer 2/3 (F1) and layer 5/6 (G1) and in the prefrontal cortex (H1). F1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.3570 for Nkx2.1Cre-cHET and p = 0.5603 for Nkx2.1Cre-cKO versus Ctrl. G1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.8491 for Nkx2.1Cre-cHET and p = 0.9244 for Nkx2.1Cre-cKO versus Ctrl. H1, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.9136 for Nkx2.1Cre-cHET and p = 0.2171 for Nkx2.1Cre-cKO versus Ctrl. F2–H2. Density of SST+ GFP+ per square micrometer in sensory cortex layer 2/3 (F2) and layer 5/6 (G2) and in the prefrontal cortex (H2). F2, One-way ANOVA with Dunnett's multiple-comparisons post hoc method, p = 0.7498 for Nkx2.1Cre-cHET and p = 0.40
留言 (0)