
記住我

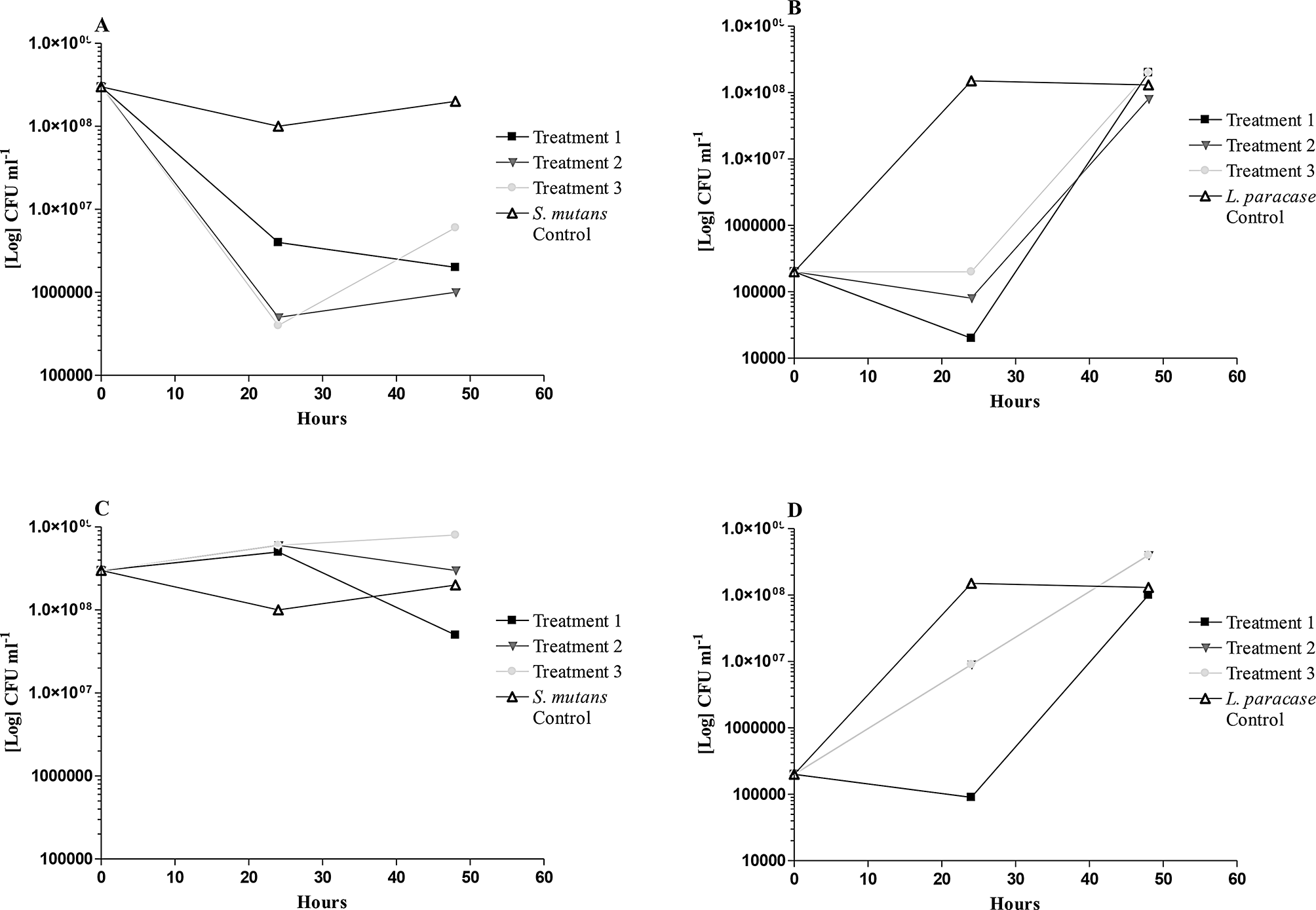





The bacterial load of S. mutans and L. paracasei were determined using colony-forming units (CFU) at the intervals of 24 and 48-hour samples respectively (Fig. 1A-D). At high concentrations tested, the bacterial load of S. mutans was reduced for all three treatments: pre- post- and simultaneous treatment after 24 h (Fig. 1A). After 48 h, a recovery of the bacterial load was observed for the post-treatment only (one-hour incubation with only bacteria), indicating that the attached bacteria could recover after initial exposure to the sample. However, with the pre-treatment (one-hour incubation with only the sample), the bacterial load continued to reduce, suggesting that the ability of the bacteria to attach and subsequently recover was reduced. For the simultaneous treatment, bacterial growth exhibited similar trends when compared with the untreated S. mutans control. At a lower concentration (half-MIC), only the pretreatment indicated a reduction in the bacterial load of S. mutans when compared to the untreated S. mutans control (Fig. 1C). At both concentrations tested, similar trends with S. mutans were seen, suggesting that the samples’ mechanism is selective for attachment and biofilm formation of the cariogenic S. mutans (Fig. 1A and D). The commensal bacteria, L. paracasei, showed an initial reduction in bacterial load at 24 h, but recovered to normal levels after 48 h, regardless of the treatment and concentration (MIC or half-MIC) used (Fig. 1B and D).
Fig. 1
The change in Streptococcus mutans and Lactobacillus paracasei bacterial load, on enamel samples, at 24 and 48 hours, after treatment (MIC and half-MIC) with the synergistic plant combination. Treatment 1: one-hour pretreatment with the sample. Treatment 2: simultaneous addition of sample and bacterial inoculum. Treatment 3: one-hour pre-incubation with bacterial inoculum. Control: untreated bacterial inoculum. (A) S. mutans at MIC; (B) L paracasei at MIC; (C) S. mutans at half-MIC; (D) L paracasei at half-MIC
The effect of the synergistic plant combination on the attachment of the bacteria to the enamel blocks under different treatments for S. mutans was investigated using FE-SEM (Figs. 2 and 3), and compared to the adherence of a commensal bacterium, L. paracasei (Fig. 4). The morphology of S. mutans was significantly altered by the synergistic plant combination at both 24 and 48-hour intervals, when compared to L. paracasei, even though both are Gram-positive microorganisms. The integrity of the cell walls of S. mutans appeared to be weakened and in some cases, cell lysis was evident after the 24 and 48-hour intervals. Cell clumping occurred with Treatments 1 and 2 (Fig. 2B-C), and substantial morphological changes were observed, when compared to the untreated control after 24 h (Fig. 2A). In Treatment 3, where bacteria were added one-hour before the synergistic plant combination, streptococci chains were formed, but tended to be short and the morphology of the cells visibly altered (Fig. 2D). After 48 h of growth, no significant numbers of streptococci were able to adhere to the enamel surface (Fig. 3B) when compared to the untreated control (Fig. 3A). The effect of all the treatments of the synergistic plant combination on commensal L. paracasei was less drastic for cell morphology and adherence. Curving of bacilli and club-like cell formation was observed (Fig. 4B-D) when compared to the untreated (negative) control (Fig. 4A).
Fig. 2
SEM micrographs of Streptococcus mutans after 24 h. (A) Untreated Control. Adhesion structures (pilli) found between the bacteria (indicated by arrows). Chain formations initiated. (B) Enamel after Treatment 1: S. mutans was added one hour after the synergistic combination at a half-MIC. Cells clumping shrinkage observed. Chain formation was affected. (C) Enamel after Treatment 2: S. mutans with the synergistic plant combination at half-MIC. Cells are clumped, and morphology distorted to the extent that individual cells are hardly recognizable (indicated by arrows). (D) Enamel after Treatment 3: S. mutans was added one hour before the synergistic plant combination at a half-MIC. Chain formation notable
Fig. 3
(A) Streptococcus mutans control after 48 h. Extensive growth and clumping of the streptococci chains can be seen, adhering to the enamel surface and each other through pellicle and glucan binding, (B) Enamel after Treatment 3, and 48 h of growth. S. mutans was added one hour before the synergistic plant combination at the MIC concentration. Chain formation is affected and the normal morphology and size of S. mutans (indicated by arrows), as seen in (A), is distorted
Fig. 4
SEM micrographs of Lactobacillus paracasei after 48 h (A) Negative control. These rods are slow-growing bacteria. Adherence to the enamel surface observed with pellicle formation (indicated by arrows). (B) Enamel after Treatment 1: L. paracasei was added one hour after the synergistic plant combination at half-MIC. The normal morphology of L. paracasei, as can be seen in (A), has become distorted (upper right arrow). Cell debris is also indicated. (C) Enamel after Treatment 2: L. paracasei was added together with the synergistic plant combination at half-MIC. The normal morphology of L. paracasei is distorted giving the rods a club-like appearance, cells of different sizes and cell debris. (D) Enamel after Treatment 3: L. paracasei was added one hour before the synergistic plant combination at half-MIC. The normal morphology of L. paracasei, as can be seen in the 48-hour control sample (A) has become distorted with long thin cells now evident (indicated by arrows)
Heteropyxis natalensis on its own is a significant deterrent in the adherence of S. mutans. It can prevent S. mutans from adhering, by forming a biofilm that restricts glucan binding, thereby inhibiting the bacteria from adhering to each other [9]. Heteropyxis natalensis, M. alternifolia and M. piperita all have 1,8-cineole present, which may permeabilize the bacterial membranes and facilitate the entry of other more active components. Melaleuca alternifolia is known to affect the bacterial membrane integrity and function, causing loss of intracellular material [10, 11].
The powerful antioxidant, epigallocatechin gallate, present within the C. sinensis extract (94%), binds directly to peptidoglycan in cell walls and induces precipitation [2, 12, 13]. Green tea inhibits the three processes involved in the pathogenesis of dental caries; namely adherence, glycocalyx formation and acid production [2, 14]. Membrane structures are also affected by epigallocatechin gallate. Streptococcus mutans seems to be particularly sensitive to epigallocatechin gallate but not L. paracasei [15, 16]. This may be because it inhibits glucosyltransferase (GTF) expression and activity of S. mutans. The suppressed GTF genes may, therefore, affect S. mutans on a transcriptional level, the initial attachment of S. mutans and all further formation of biofilms [15].
In another study, Bacillus subtilis cells were exposed to M. piperita essential oil at a concentration of 1.13 mg ml− 1 for 4 h and examined using scanning electron microscopy (SEM) [17]. The sample had considerable morphological alterations when compared to B. subtilus control - described as intact, rod-shaped, separated from each other, turgid and whole with a smooth surface while the M. piperita oil treated cells appeared to be partially deformed with frequent depressions on the cell surface. This was ascribed to the terpenes; menthone; isomenthone; 1,8-cineole; menthyl acetate; menthofuran; limonene; β-myrcene; β-caryophyllene; pulegone and carvone, present in the essential oil [18, 19]. The same authors noted that certain terpenes may increase membrane fluidity and alter the membrane permeability, resulting in alteration of membrane properties and functionality [17].
Our results corroborate those of Tyagi and Malik [17], where similar morphological changes were observed in S. mutans in Fig. 2 (B-C) and in L. paracasei Fig. 4 (C) where the bacteria appeared to be partially deformed and displayed loss of turgidity and separation.
In a study by Cho, et al., (2010), polyphenols extracted from green tea (C. sinensis) were evaluated for their antimicrobial effects and inhibition of biofilm formation properties against twelve oral microorganisms which included: Streptococcus mutans, S. sanguis, S. sorbrinus, S. mitis, S salivarius, Lactobacillus acidophilus, L. plantarum and Candida albicans. Tea polyphenols inhibited the growth of all the tested microorganisms at 2 mg ml− 1 within 5 min. Depending on the bacteria, various morphological changes, such as the presence of perforations, formation of cell aggregates, and leakage of cytoplasmic materials were also observed by SEM. Biofilm formation of S. mutans and S. sanguis were inhibited on teeth [20], but no indication was given on maintaining a level of resident microbiota on the teeth. Other plants have also been investigated for their abilities to prevent the adherence of S. mutans. Saussurea lappa (commonly known as costus), has traditionally been used for the treatment of halitosis, dental caries, and periodontal disease.
The effect of S. lappa on adherence to saliva-coated hydroxyapatite beads (S-HAs) by S. mutans (ATCC 25175) was indicated when the ethanolic extract (0.25-4 mg ml− 1) significantly reduced the adherence of S. mutans to S-HAs. Saussurea lappa may also reduce the cell surface hydrophobicity of S. mutans, which is an important requirement for the bacterium to adhere to the tooth surface [21].
A general toxicity test was conducted on U937 cells for both the synergistic plant combination and each component. The relative concentrations of the individual components in the synergistic plant combination are provided in Table 2. According to Hussain et al., [22], the toxicity of a tested sample can be categorized based on its IC50 values where: an IC50 < 10 µg ml− 1 represents high toxicity; IC50 values between 10 and 100 µg ml− 1 represents potential toxicity; IC50 values between 100 and 1000 µg ml− 1, potentially harmful and an IC50 > 1000 µg ml− 1 represents non-toxic. According to the classification, H. natalensis would be considered potentially toxic (35.56 ± 0.16 µg ml− 1) while C. sinensis and M. alternifolia would be categorized as non-toxic. Mentha piperita exhibited an antiproliferative effect at 0.015% v/v. It is, however, interesting to note that when H. natalensis was combined with C. sinensis and the essential oils in the synergistic composition, the IC50 value almost doubled (> 62.5 µg ml− 1). This may be indicative of a cellular protective function for the macrophage U937 cells, induced by the combination. Wagner [23], states that this may be due to the synergistic effect of nullifying toxicity or adversely acting substances by a single component added to the extract.
留言 (0)