
記住我






On the three days following the last habituation session, animals were exposed to three different manipulations to habituate them for the IMO. These manipulations were given once per day. First, animals were exposed for 20 min to a new environment consisting of a box with an area of 25 cm2 for free exploration. The following day, animals were introduced inside a 50 mL Falcon tube, with a drilled cap to permit air transpiration and facilitate breathing [23]. The day after these habituation sessions, animals were exposed to IMO for 2 hours. The IMO procedure was conducted in a room separated from housing and behavioral tests. Each animal was immobilized by gently attaching their four limbs in a prone position to metal arms attached to a plastic board restricting their movement, as previously described [11]. Recordings of calcium activity and behavior were taken for the first and the last 10 min of the IMO procedure. After the exposure to IMO, animals were left undisturbed for six days until the FC session.
Fear conditioning and extinctionThe protocol used for FC six days after IMO was performed in context A and consisted of a 5-minute intra-session habituation; five 30-second-long tones (30s, 6 kHz, 75 dB) coterminated with an electric footshock (1s, 0.3 mA), and separated by 180-second intertrial intervals (ITI); and 180 final seconds after the last CS-US pairing. The two following days, animals underwent two FE sessions (one session per day), performed in context B. Each FE session consisted of a 5-minute intra-session habituation followed by 15 uncoupled (no shock) 30-second-long tones (30s, 6 kHz, 75 dB), spaced by 30-second ITI (Fig. 1C), finishing with 30 s after the last tone.
Calcium imaging recording and analysisGeneral considerationsThroughout sessions, calcium activity (GCaMP6f 488 nm fluorescence) was registered using a Miniscope (v3.2 UCLA University, CA, USA) coupled to the Miniscope DAQ hardware and software. The Miniscope system is an open-source microscopy platform for recording and analyzing neural activity in freely behaving animals, developed by Daniel Aharoni Lab (UCLA University, CA, USA).
The Miniscope relied on a 0.25 pitch, 2 mm-diameter GRIN lens (Inscopix, CA, USA), coated with black heat shrink tubing. The Miniscope DAQ software was set up with a recording rate of 30 frames per second. The LED intensity and digital gain were adjusted for each animal and kept constant throughout the experiment [24].
For FC, data were obtained from 5 females and 6 males; for FE1, we have 5 females and 6 males; and for FE2, there are 5 females and 6 males for all calcium imaging recordings. Exclusion from one session was due to outlier removal or recording issues. It did not lead to the removal of the animals from all tasks given that the experiment was performed on all animals using the same conditions. However, animals exhibiting recording issues either at the start or end of the IMO were excluded from all IMO analyses, leaving 5 males and 5 females in this task.
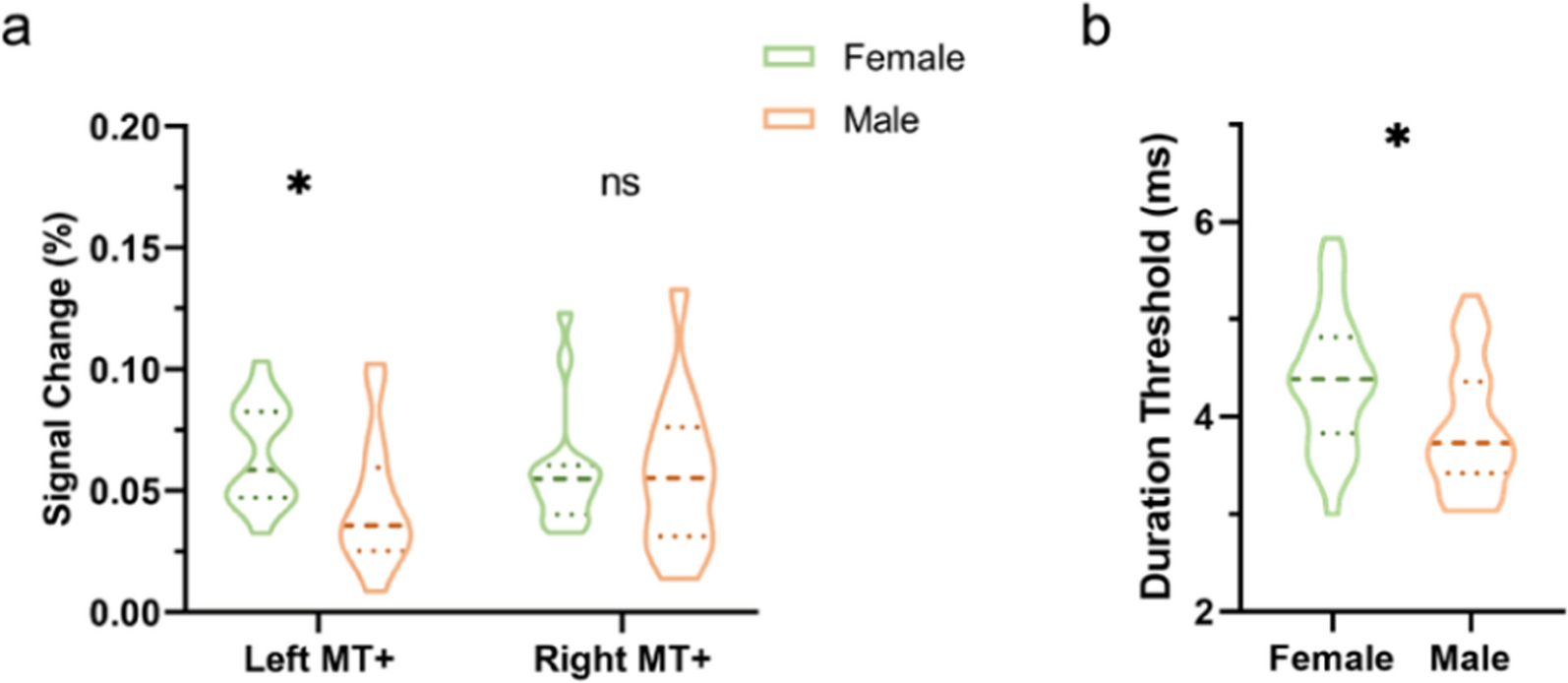
Calcium signal deconvolutionWe used the publicly available algorithm, Constrained Non-Negative Matrix Factorisation optimized for Endoscope signals (CNMF-E), to deconvolve the calcium signal [25]. CNMF-E first identifies non-linear shifts in the visual field using an optimized template-matching algorithm, NoRMCorre [26]. All data are reconstructed using the extracted shift vectors to erase motion artifacts. Following this motion correction, the visual field is segmented into regions of interest (ROI) using the CNMF-E algorithm. This delineates the spatial footprints of single-neuron somas. By measuring the fluorescence pattern of different ROI, CNMF-E can identify and discriminate different, yet overlapping, neurons and demix their signal where they overlap (“Trace demixing”, Fig. 2). Lastly, each ROI’s fluorescence trace is fit by a spike-decay model of the calcium imaging indicator and transformed to express fluorescence as a relative change in fluorescence, \(\:\varDelta\:F\) (Fig. 2A).
Fig. 2
Computational methods for the calcium imaging videos. (A) shows the overall working pipeline of CNMF-E from the original video until the extraction of filtered traces. From left to right are shown an unprocessed frame from an IMO session, the spatial footprints for the set session, and the raw and filtered traces for the first 15 identified ROIs. (B) shows, for a single session, the average shuffled ϕ coefficient for all the neurons in a single session with their 95 Confidence Interval (CI). In red are shown the ϕ scores for the original traces. Dots label all neurons exhibiting a ϕ score outside the 95CI range. (C) shows sample stimulus-excited (red) and stimulus-inhibited traces (blue)
For all sessions, the following set-up was used: spatial downsampling = 3; motion correction = non-rigid; temporal downsampling = 3; dendrite identification = false; spatial algorithm = hals; include residuals = false; deconvolution method = foopsi, minimum spike size = 5 x noise; de-trend method = spline; background model = svd.
For fear conditioning (FC) and fear extinction (FE1 and FE2), all single-neuron traces were obtained by the deconvolution step. Furthermore, they were averaged and expressed as a scaled difference from baseline fluorescence levels for whole-field analyses (\(\:\varDelta\:F/_\)), where \(\:_\) is the median of the complete fluorescence, and \(\:\varDelta\:F\) is the difference between \(\:_\) and the fluorescence trace. We chose the median as the baseline given the vulnerability of the mean to transient changes in the signal. This procedure was carried out to extract, from the values of fluorescence in the tones, the global fluorescence baseline. The goal was to compare data in the tones without biases. For IMO, this step was not necessary, due to the stressful nature of the procedure, which was constant throughout the two hours of the process. Hence, it was possible to analyze fluorescence traces without impactful biases from external inputs (ex.: the shock, or the sound of the tone), which make it necessary to apply the previously described normalization step.
PL calcium activity during IMOThe movement of the animal is the main behavior that can be registered while it is immobilized. This movement can be interpreted as the attempt or struggle of the animal to free itself from the IMO. We quantified the movement of the animals as the frequency and the length of high-activity epochs common during the IMO while simultaneously recording PL calcium activity with the Miniscopes. We developed a MatLab code to compute global movements within the field of vision. All RGB frames were converted to grayscale, and edges were extracted using the Sobel edge detection method (built-in MATLAB® function). Thresholds for edge detection were extracted on a frame-to-frame basis using the canny method [27] and scaled by a fudge factor of 0.5. This allowed us to reduce the amount of data processed by avoiding taking into consideration the entirety of the animal. This edge detection returned, for each frame, a binary edge mask. We expressed this mask as a width \(\:\times\:\:\) height matrix. The movement was characterized as the percentage of pixels at t = n, that were not present in t = n−1. These scores were divided by the count of total edge-bearing pixels identified, t = n. Using test videos, we validated that this measure is a practical approach to detect movement at the field level. Given that trial-to-trial changes in camera position, environment, and mouse dimensions can impact this variable, all movement traces were shifted to a median equal to 0 and scaled by the 95% confidence interval.
The movement score was used to identify periods of high activity of the animal during the IMO. By measuring the movement in the video field, we found that their distribution can be best described by two distinct normal distributions. These correlate to low and high movement frames. We defined the intersection between the two distributions as the classifier to select high-movement frames. For each animal, we calculated the number of frames that animals spend in this high-movement status. With this method, we analyzed differences in the percentage of high-movement events, in their length and frequency.
Furthermore, we decided to test calcium activity and movement for correlation, employing Pearson’s Test. First, we analyzed the data globally, by testing movement, during early and late IMO, with the global fluorescence activity in both females and males (as two different groups). Secondly, we decided to look into individual neurons and identify the specific percentages of correlated (positively and negatively) and uncorrelated neurons. We use Pearson’s correlation test between individual neuronal fluorescence traces and movement scores for each animal in early and late IMO. We defined neurons as uncorrelated with p > 0.05, positively correlated with p < 0.05 and R factor > 0, and negatively correlated with p < 0.05 and R factor < 0. We divided the neurons into two groups, belonging to females and males, and the analysis was done for early and late IMO.
Fear memory, freezing, and PL calcium activityFor FC and FE sessions, behavioral video data was obtained with the Freezing v1.3.04 software (Harvard-Panlab-Barcelona, Spain), with a resolution of one immobility measure per second (expressed in %). Data were binarized as freezing or non-freezing. The first is regarded as a percentage of immobility beyond a respiratory rate above 0.7.
We decided to test calcium activity and freezing for correlation, employing Pearson’s Test. First, we analyzed the data globally, by testing global fluorescence and freezing during a specific session (FC, FE1, and FE2) in both females and males (as two different groups). Secondly, we decided to look into individual neurons and identify the specific percentages of correlated (positively and negatively) and uncorrelated neurons. We use Pearson’s correlation test between individual neuronal fluorescence traces and freezing for each animal in FC, FE1, and FE2. We defined neurons as uncorrelated with a p > 0.05 significance, positively correlated with p < 0.05 and R factor > 0, and negatively correlated with p < 0.05 and R factor < 0. We divided the neurons into two groups, belonging to females and males, and the analysis was done for all sessions (FC, FE1, and FE2).
Identifying stimulus-responding neuronsCurrent methods for estimating neuronal activity in fear memory with Miniscopes often rely on global-field measurements. However, this approach can result in the simplification of a population with heterogeneous neuronal responses. To address this issue, we developed a different method to compute the modulatory effects of the different tones on calcium dynamics. Firstly, we defined a calcium event as a local maximum in the filtered calcium trace. Following, each neuron’s activity was expressed as a Boolean (true or false) indicating the temporal location of calcium events. For each neuron, the Phi Coefficient (ϕ; a measure of association for two binary variables), was computed to describe the association between tone presentation and calcium responses. To this purpose, we define “true positive” as a calcium event during tone presentation and “false negative” as a frame without a calcium event during tone presentation.
To avoid delimiting a global ϕ threshold, all event matrices were randomized, and ϕ scores were computed 1000 times. Results were stored in a matrix, R. Original scores were expressed as Z distance from the population matrix R. Z distance (or absolute Z-score) is a statistical measure that quantifies the distance between a data point and the mean of a dataset.
All formulas used were included in Supplementary Material.
Histological processingAfter termination of the protocols, animals were anesthetized with Isoflurane, perfused first with saline serum (0.9% NaCl) and afterwards with 4% paraformaldehide (Casa Alvarez, Spain) for later histological processing and validation of AAV injection and lens placement (Fig. 1D, Fig. S1). Brains were extracted, cryopreserved in 30% Sucrose and frozen in dry ice-cooled (-55oC) isopentane (Sigma-Aldrich). Then, 30 μm coronal sections of the PL were obtained using a cryostat (Leica CM3050). Sections were then stained with DAPI (1:20000), and mounted on Superfrost slides (Thermo Scientific, USA). Next, a fluorescence microscope (Nikon Eclipse 90i) was employed to capture images depicting the distribution of the vector-associated reporter (EGFP). These images were then overlapped with the DAPI image (Fig. 1D).
Statistical analysesWe used MATLAB code in the R environment including the following tests: Shapiro-Wilk for testing for violations of normality within groups and across the global sample. Levene’s tests for equality of variances. Grubb’s recurrent test for outlier identification. One-way and repeated measures ANOVA for testing for differences between and within groups. We used Tukey’s test for multiple comparison analysis. Furthermore, to test for a correlation between calcium fluorescence and either freezing or motion, we used the Pearson’s Correlation Test.
Statistical analyses were integrated with the MATLAB code and tailored to the specific data tested. All codes (MATLAB and R) of the complete pipeline and all the hyperparameters are available on our GitHub.
留言 (0)