
記住我

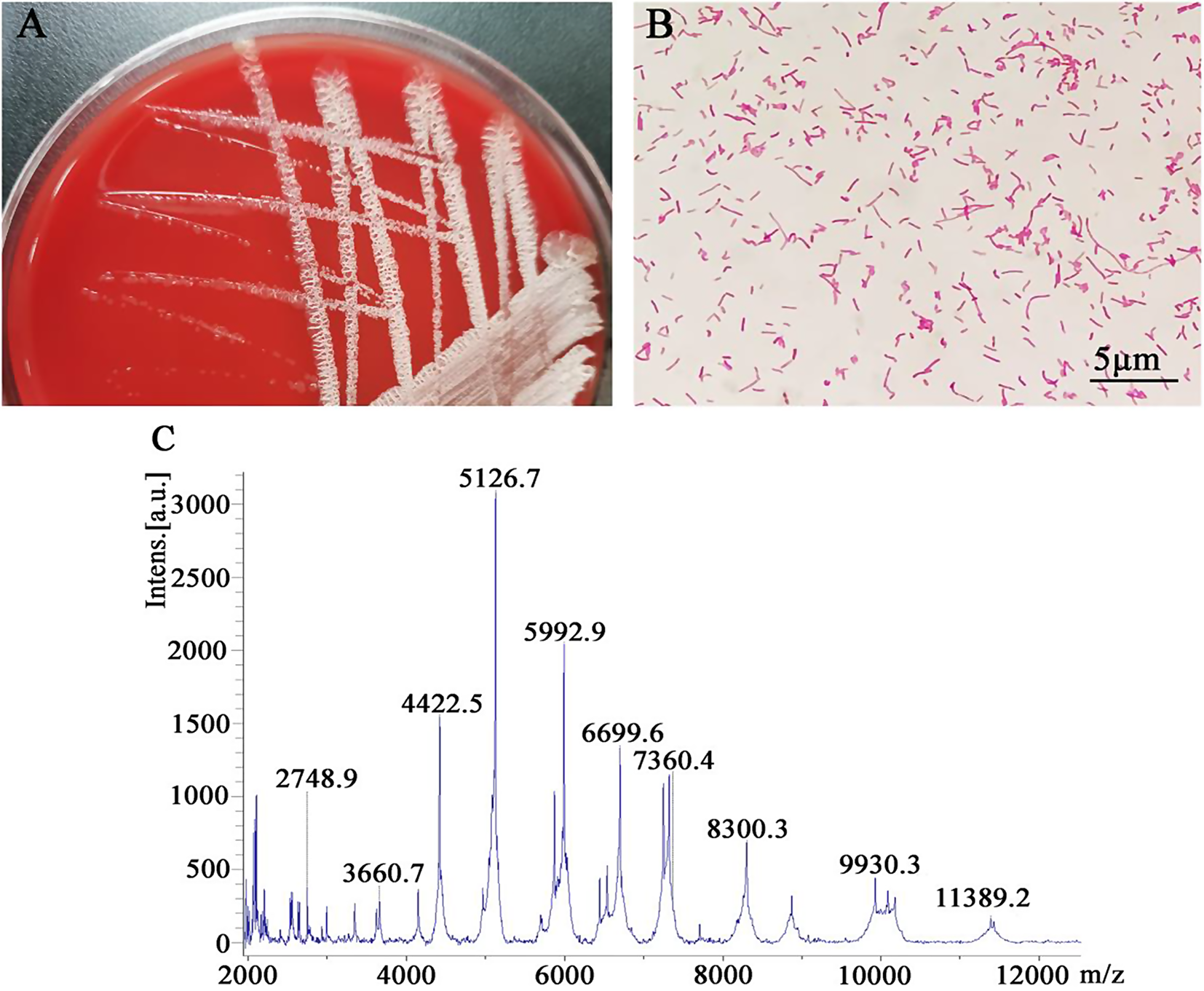






The study analyzed 78 MDR P. aeruginosa isolates from various clinical specimens, finding no significant association between gender and clinical specimens, despite a significant gender-related association (p > 0.05) (Table 1).
Table 1 Epidemiological and clinical characteristics of the Pseudomonas aeruginosa isolatesAmong the discovered P. aeruginosa isolates, the majority (60.3%) were from females, while 39.7% were from men. The highest infection rate was observed among individuals aged 60–80 years (51.2%). This result aligns with a prior investigation conducted by Subramaniyan and Sundaram, which revealed that 31.1% of the samples were male and 68.8% were female [31]. The majority of isolates (35, 44.8%) were obtained from sputum (35, 44.8%), followed by urine samples (18, 23%), swabs (15, 19.2%), cerebrospinal fluid (8, 10.2%), and blood (2, 2.5%) (Table 1). The largest number of clinical isolates was obtained from sputum, indicating the presence of respiratory tract diseases. The findings align with the research conducted by others, who similarly observed a significant occurrence of P. aeruginosa in tracheal aspirates [31, 32].
Antimicrobial resistanceThe Vitek-2 automated system test indicated that all 78 P. aeruginosa isolates exhibited significant resistance to the antimicrobials that were evaluated. A total of 13 antibiotics belonging to seven different classes were chosen for antibiogram profiling because of their widespread usage in hospitals as medications to combat pseudomonas infections. The isolates shown substantial resistance to multiple tested antimicrobial drugs (p < 0.001) (Table 2).
Table 2 The antimicrobial resistance profile of Pseudomonas aeruginosa isolates. The results are repeated triplicate p < 0.001The study revealed that the greatest level of resistance was observed against β-lactam antibiotics, specifically within the penicillin class [piperacillin 63 (80.7%), piperacillin/tazobactam 61 (78.2%)], cephalosporin class [cefepime 57 (73%), ceftazidime 53 (67.9%), and cefoxitin 54 (69.2%)], as well as the tetracycline class [Tigecycline 50 (64.1%)]. Table 2. These findings are consistent with prior research [1, 3, 20]. The increasing prevalence of drug resistance poses a significant challenge for treatment options, particularly with the emergence of high carbapenem resistance among Gram-negative bacilli isolates, which has been extensively reported in various countries. Consequently, carbapenem resistance has become a critical global public health crisis [33]. In the present study, the rate of imipenem resistance against MDR-P. aeruginosa was moderately high at 51.2% (Table 2). This emergence of such resistant isolates is a cause for concern in public health. A study in Isfahan found that out of 106 P. aeruginosa isolates, 62 (58.5%) were resistant to imipenem [34]. This is lower than previous reports, which reported 17.7% resistance [1]. In Iran, imipenem resistance in burn and non-burn patients was 83.2% and 57.5%, respectively [35]. Additionally, 33.3% of isolates showed resistance to meropenem, a highly effective antipseudomonal antibiotic. This finding aligns with studies in Bangladesh and Nepal [20, 36]. Resistance to other classes of antibiotics was linked to resistance against carbapenem antibiotics. Rates of resistance were found to be tigecycline (64.1%), trimethoprim/sulphamethoxazole (62.8%), levofloxacin (38.4%), ciprofloxacin (35.8%), gentamicin (34.6%), and amikacin (33.3%). Interestingly, meropenem and amikacin were effective against approximately 70% of the MDR-P. aeruginosa isolates. Amikacin was found to be the most effective antipseudomonal antibiotic for MDR-P. aeruginosa infections and a suitable treatment option for carbapenem-resistant P. aeruginosa isolates. Amikacin's resistance rate (33.3%) aligns with de Sousa et al.'s findings [2], but a different study reported a high resistance rate (92.4%) in burn infections caused by carbapenem-resistant P. aeruginosa isolates [37]. In brief, 85.8% of strains exhibited resistance to at least three classes of antimicrobials, making them MDR-P. aeruginosa. This result is consistent with the high rates of MDR-P. aeruginosa isolates in Iran, where 89.4% of the isolates were found to be MDR [38]. There is a growing occurrence of MDR-P. aeruginosa isolates worldwide, and this study has found higher rates than prior findings. Research has also indicated that patients infected with P. aeruginosa and Acinetobacter spp. have a significant risk of death, with mortality rates ranging from 40 to 65% [39]. The high resistance to carbapenem antibiotics is concerning from a public health perspective, as these antibiotics are essential for treating infections when lower-class antibiotics are no longer effective [2]. The higher incidence of multidrug-resistant (MDR) patterns among strains in this study may be attributed to factors such as self-medication, empirical usage, and excessive use of carbapenems and third-generation cephalosporins. Additional surveillance initiatives are required in this region to effectively tackle the escalating issue of carbapenem resistance [40].
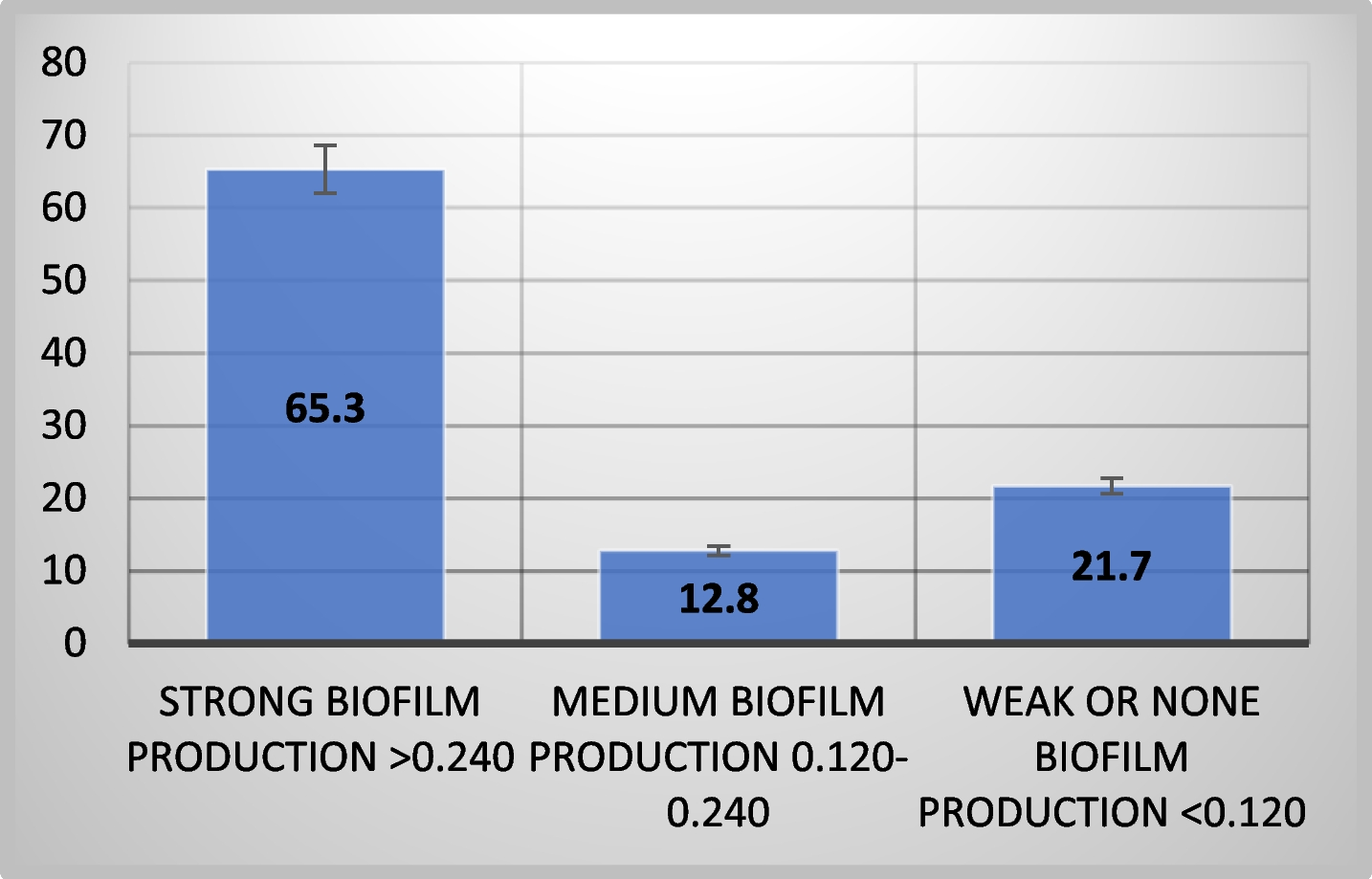
Biofilm production assayThe study revealed that 78.2% of 78 isolates were biofilm producers, indicating their higher resistance to antibiotics compared to free-floating cells [9].
Figure 1 shows that most strains (65.3%) had strong biofilm formation capacity, while 12.8% and 21.7% exhibited moderate or weak biofilm-forming potential, respectively, using the tissue culture plate method.
Fig. 1
Biofilm-production capacities among Pseudomonas aeruginosa isolates
Most pathogenic P. aeruginosa strains can form biofilms, making them difficult to remove from hospital environments and medical equipment. There was a significant (p < 0.0001) difference in biofilm production abilities between tested MDR-P. aeruginosa isolates. The study found that all P. aeruginosa strains, including clinical specimens, can form biofilms under laboratory conditions, although the extent of biofilm formation varies. This aligns with previous studies, where 19.87% of isolates were weak/non-biofilm producers, 20.86% were moderate producers, and 59.27% were strong biofilm producers [3]. In contrast, de Sousa et al. reported higher biofilm production rates, with 92% of P. aeruginosa isolates producing biofilms [2].
Prevalence of carbapenemase genesThe MDR profile, increased incidence, and rapid dissemination of carbapenemases across species through transmissible genetic factors make them the most urgent resistance problem in Gram-negative bacteria [41, 42]. These genes often carry other antimicrobial resistance genes, leading to MDR or extensively drug-resistant traits, limiting treatment options and posing a triple threat. Figure 2 displays the prevalence of carbapenemase genes in 30 isolates that were tested. A study found that all 30 P. aeruginosa isolates tested had one or more carbapenemase genes. The most common carbapenemase gene was blaOXA-48, present in 83.3% of isolates. Other common carbapenemase genes included blaNDM (66.6%), blaIMP (50%), and blaVIM (33.3%), and none of the isolates tested positive for blaKPC (Fig. 3 A, B, C, and D). 76.6% of positive isolates had multiple carbapenemase genes, ranging from two to four. Statistical analysis showed a significant difference (p < 0.001) in carbapenemase resistance genes prevalence among MDR-P. aeruginosa isolates.
Fig. 2
Frequencies and distribution of carbapenemase genes in P. aeruginosa isolates
Fig. 3
Detection of carbapenemase genes IMP, and VIM in carbapenemase-producing P. aeruginosa isolates. Lane M represents a 1-kb DNA ladder and lane C is a negative control. A The gel electrophoresis displayed the presence of carbapenemase gene NDM (621 bp). B Lanes 1–5 represent the positive IMP carbapenemase gene. C Gel electrophoresis revealed the presence of blaVIM (390 bp). D The gel electrophoresis showed the presence of the carbapenemase gene blaOXA-48 (438 bp)
This study found that blaOXA-48 is the most common carbapenemase gene, followed by blaNDM. Subramaniyan and Meenakshi's found that 13.1% of MBL-producing P. aeruginosa strains from the ICU were 13.1% blaVIM-4, 9.8% blaVIM–5, and 3.2% blaVIM–38, with none testing positive for KPC, NDM, and IMP genes [31]. Sheikh et al. identified 236 carbapenem-resistant P. aeruginosa isolates, with 116 carrying MBL genes and 29 testing positive for blaNDM-1 [35]. Similarly, Ramadan et al. found that genes encoding VIM, GES, NDM, and IMP were detected in 50%, 40.9%, 27.3%, and 18.2% of isolates [8]. None of the tested isolates showed the presence of the blaKPC gene, as indicated by earlier research findings [8, 35]. This could be attributed to the existence of additional class A carbapenemases [43]. OXA-48, a highly prevalent worldwide ailment, has reached epidemic proportions in multiple Mediterranean nations, such as Turkey, Iran, Morocco, and Lebanon [35, 44]. The study found that a significant proportion of isolates (76.6%) carried multiple carbapenemase genes, indicating their ability to produce MDR determinants. This is a major concern for antibacterial treatment strategies, as it has been reported in other studies [1, 35].
Biosynthesis of TiO2NPsThe study examined all isolates for TiO2NP production, but only one P. aeruginosa isolate demonstrated the ability to synthesize TiO2NPs.
Characterization of TiO2NPs:1-visual observation and UV–visible analysis
The stability and formation of synthesized TiO2NPs in an aqueous colloidal solution were determined through visual observation and UV–visible analysis [21]. The isolated P. aeruginosa strain successfully biosynthesised TiO2NPs by exposing bacterial cultures to TiO2, which was confirmed by the color changes from yellow to white precipitate, indicating the synthesis of TiO2NPs (Fig. 4 A). The color generation in TiO2NPs is due to the excitation of surface plasmon in metal nanoparticles [45]. UV absorption analysis confirmed the synthesis, with a clear absorption peak indicating the anatase phase of nano-TiO2 (Fig. 4 B). The cut-off wavelength was 316 nm, consistent with previous research on TiO2NP synthesis using various bacterial strains.
Fig. 4
The positive result of color change and UV–visible spectroscopy of biosynthesized TiO2NPs (A and B), XRD results (C), FT-IR spectra analysis (D), and EDX analysis (E)
Microbial systems, including bacteria, are efficient in producing reducing components for nanomaterial synthesis due to richness in several biomolecules can transform the Ti salts to TiO2NPs [46]. P. aeruginosa was chosen for TiO2 synthesis due to its environmental compatibility, low energy consumption, and cost-effectiveness [47].
2- X-ray diffraction analysis
The synthesized nanoparticles' chemical composition and crystalline properties were verified using X-ray diffraction (XRD) analysis [21]. The crystal structure of TiO2NPs was examined using an X-ray diffractometer, which confirmed the formation of TiO2NPs from P. aeruginosa culture supernatant, with XRD patterns matching standard diffraction data (JCPDS card number: 98–017-2916). The XRD analysis of synthesized TiO2NPs revealed distinct diffraction peaks at various angles, including 25.30°, 36.96°, 37.81°, 38.56°, 48.04°, 53.88°, 55.07°, 62.68°, 70.31°, 75.09°, and 76.09°, corresponding to the crystal planes 101, 103, 004, 112, 200, 105, 211, 204, 220, 215, and 224, respectively (Fig. 4 C). The synthesized sample has a small size, crystalline structure, and high purity, as indicated by sharp diffraction patterns [26]. The XRD results from Fig. 4 C indicate that the nanoparticle structure primarily consists of the anatase crystalline phase, known for its high photocatalytic activity, and the main peaks' positions align with previous literature findings [26, 48].
3-Fourier Transform Infrared Spectrometry (FTIR)
Fourier transform infrared (FTIR) spectroscopic analysis was conducted to investigate the interaction between the NPs and capping agents. The FTIR spectra were recorded in the wavenumber range of 400–4000 cm-1. The FTIR spectra of TiO2NPs synthesized by P. aeruginosa exhibited prominent peaks at 3258.14, 2988.16, 2901.38, 1636.3, 1540.85, 1394.28, 1250.61, 1055.84, and 473.43 cm − 1 (Fig. 4 D). The biosynthesized TiO2NPs showed a broad band at 3258 cm − 1, indicating O–H stretching due to the alcoholic group. The peaks at 2988 cm − 1 and 2901 cm − 1 corresponded to the C–H of CH2 and CH3 groups in aliphatic chains, while signals at 1636 cm − 1 and 1540 cm − 1 indicated the presence of amide and amine groups.
The FTIR analysis was used to investigate the reduction of TiO2NPs by biomolecules in microbial cells. The peaks at 1394 cm − 1 and 1250 cm − 1 were attributed to C-O stretch vibrations, possibly indicating an alcohol or carboxylic acid group. The bands at 1055 cm − 1 and 473 cm − 1 represented C-O stretching vibrations of aliphatic amines and Ti–O stretching vibrations respectively. These biomolecules may also play a role in nucleation and biosynthesis processes, acting as stabilizing and capping agents [49, 50].
4- Energy-dispersive spectroscopic analysis (EDX)
The energy-dispersive spectroscopic analysis confirmed the elemental distribution of Ti and O elements, confirming the formation of TiO2 (Fig. 4 E), in line with previous studies.
5- Morphology and shape of TiO2NPs
1-Transmission electron microscopy (TEM)
The study analyzed P. aeruginosa nanoparticles using TEM to determine their size, morphology, and distribution at the nanoscale. The TiO2NPs were found to be predominantly spherical and ellipsoidal, with uneven distribution Fig. 5 A. The average diameters and size distributions were determined using ImageJ software, with the majority falling within the 30–40 nm range, accounting for 18% of the distribution (Fig. 5 C). The study's findings, compared to previous studies by Khan, Fulekar, and Eisa et al. [22, 51], were found to align with the literature, with a size distribution histogram indicating sample polydispersity, a common characteristic of nanoparticles synthesized through biosynthetic methods [52]. The study found that most TiO2NPs were spherical or round to ellipsoidal in shape, with both individual NPs and agglomerates observed (Fig. 5 A). The nanoparticles appeared separated from each other, suggesting spiraling peptides around them.
Fig. 5
(A) TEM analysis and (B) FE-SEM analysis of biosynthesized TiO2NPs (C) Histogram of the particle size distribution of the biosynthesized TiO2NPs
2- Field emission scanning electron microscopy (FESEM)
The surface morphology and shape of synthesized TiO2NPs were analyzed using SEM, with the results showing uniform spherical shapes and dimensions. This aligns with previous studies [26, 28]. Additionally, the results of Mathesh et al. showed that Spirulina-mediated TiO2 nanoparticles were efficient against multidrug-resistant bacteria due to their spherical form with aggregation in SEM imaging and XRD analysis, which showed 61.4% crystallinity with anatase phase [53]. The SEM images are shown in Fig. 5 B.
The antibacterial efficacy of TiO2NPsTable 3 presents the disc diffusion method results for evaluating the effects of TiO2NPs with and without different β-lactam antibiotic combinations, on MDR strains of E. coli, Klebsiella sp., A. baumannii, Proteus sp., and P. aeruginosa (Table 3). The bacteria were tested against four antibiotics: imipenem, ceftazidime, piperacillin, and cefoxitin. The results showed that all tested antibiotics were ineffective against the bacteria, which exhibited resistance to these antibiotics, according to the Clinical and Laboratory Standards Institute (2020) (Table 3 and Fig. 6 C).
Table 3 Mean inhibition (mm) and fold area increase of different antibiotics, TiO2NPs, and combined against MDR bacteria. The test is repeated 3 times. *p < 0.01Fig. 6
The combined effect between antibiotics and biosynthesized TiO2NPs against Proteus sp. (A), Acinetobacter baumannii (B), and Antibiotic resistance in Proteus sp. (C)
The biosynthesized TiO2NPs, at a concentration of 5000 μg/mL, demonstrated a significant (P < 0.01) antibacterial effect against MDR A. baumannii and Proteus sp. strains. The study found that A. baumannii and Proteus sp. showed the largest zone of inhibition (22 mm and 21 mm, respectively), while E. coli, Klebsiella sp., and P. aeruginosa had zones of inhibition of 18 mm, 20 mm, and 19 mm, respectively. The study found that TiO2NPs were more effective against MDR Gram-negative bacilli strains than all tested antibiotics due to their resistance to antibiotic discs (Table 3 and Fig. 6 A and B).
TiO2NPs are a type of metal oxide nanoparticles that have been extensively studied for their antimicrobial properties. They have been found to possess the potential to eradicate both Gram-positive and Gram-negative bacteria [16, 45]. When compared to antibiotics alone, TiO2NPs demonstrated the strongest antibacterial efficacy against manty pathogens [54]. Studies have shown that P. aeruginosa synthesizes TiO2NPs with antibacterial activity, and similar findings have been observed against other bacteria [51, 55,56,57]. A study in Iraq [58] found that TiO2NPs at a concentration of 500 μg/mL showed the highest zone of inhibition against MDR test organisms, with a maximum of 24 mm against Streptococcus pyogenes, showing high sensitivity even at 31.25 μg/mL. Proteus vulgaris showed the least sensitivity. Gram-negative bacteria have higher negative charges and stronger adhesion to positive surfaces, making biosynthesized TiO2NPs more effective against them, regardless of resistance level. It is possible that oxidation and cell death are brought on by the electromagnetic attraction between bacteria and metal oxides. [59]. The bactericidal effect of tested nanoparticles may also be due to reactive oxygen species (ROS), primarily hydroxyl radicals (OH), decomposing bacterial outer membranes, leading to phospholipid peroxidation and oxidative cell death [51]. In addition to rupturing cell membranes, the main role of cytoplasmic ROS generation in NPs' antibacterial actions is to seriously damage bacteria's DNA, which eventually results in the bacterial demise. Furthermore, interactions between NPs and phosphate components within the bacterial cytoplasm may result in the formation of persistent complexes that disrupt essential bacterial enzymes [54, 60].
留言 (0)