Anatomy
The sgC has been identified in the Göttingen minipig displaying a similar topography and cytoarchitecture as the human subgenual region (Brodmann 1909). Our findings are in accordance with available publications on the frontal cortex and cingulate gyrus of human, non-human primate, rat and swine (Barbas and Pandya 1989; Carmichael and Price 1994; Jelsing et al. 2006; Vogt et al. 1986; Vogt and Pandya 1987; Vogt et al. 1987; Vogt et al. 1995; Brodmann 1909; Stephan 1951; Walker, 1940; Bjarkam et al. 2017a).
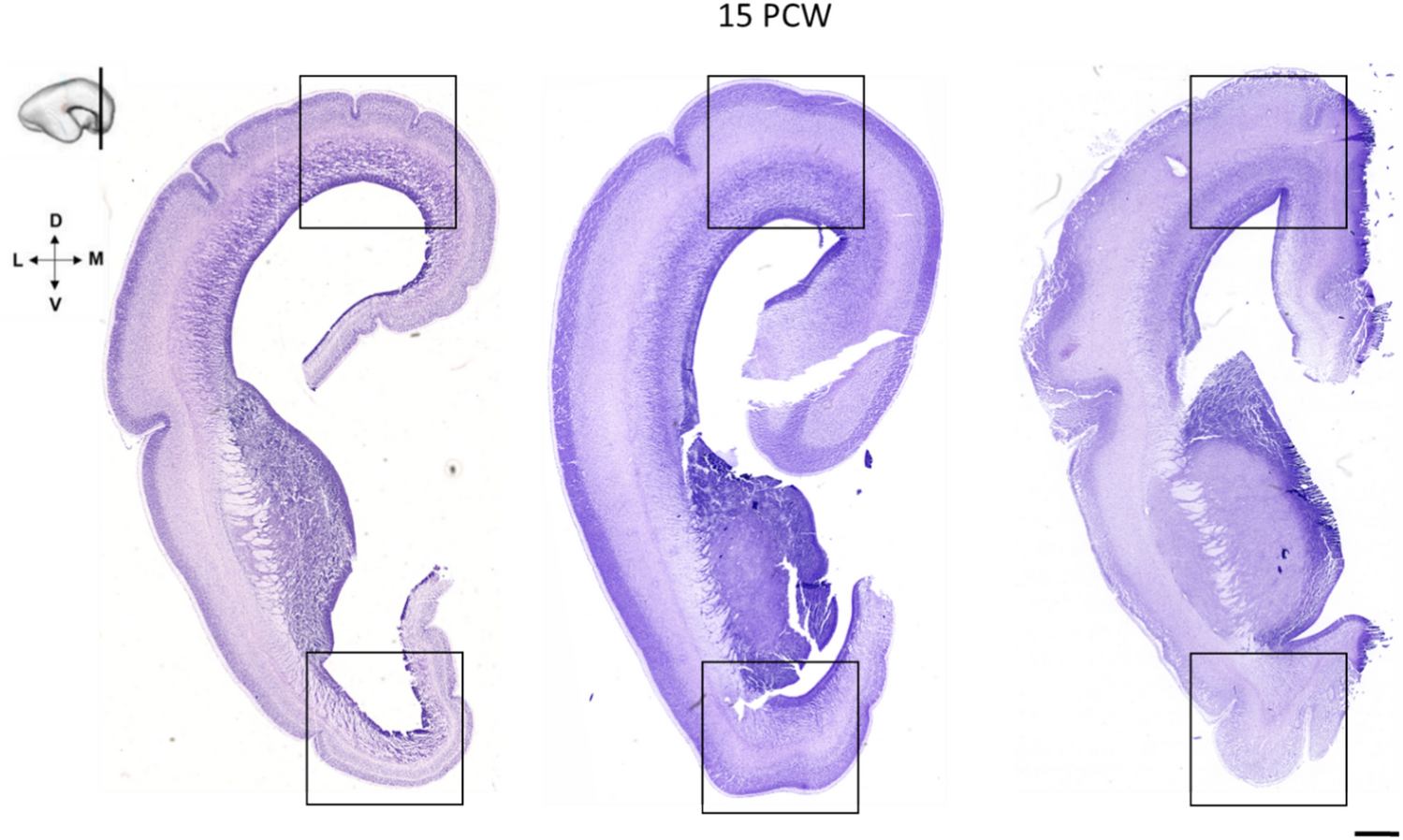
The Göttingen minipig BA25 is agranular cortex characterized by cell-poor cortical columns, located perpendicular to the pial surface, which separate the otherwise cell-rich cortex of layer II and III (Fig. 5). In layer V the cells are more evenly distributed but still of similar size as seen in layer III, while layer VI contains more widely dispersed neurons (Fig. 5). The human area 25 is likewise characterized by such cell-poor columns. It also displays a definite layer II with small aggregation of neurons and a layer VI with small neurons. However, there is a subdivision of layer V into Va and Vb and a more apparent distinction between layer III and V in contrast to what we found in the minipig (Vogt et al. 1995, 2004). Vogt et al. further divided area 25 into a rostral area 25, dissociating from a less differentiated caudal area 25 similar to the area 25 defined by Brodmann. A similar change towards less cortical thickness and layering and more homogeneous cells is noticed when moving posteriorly through the minipig area 25 towards BA33 (Figs. 3, 4, 5). In an extensive investigation on the macaque monkey prefrontal cortex, Walker (1940) identified an area 25 situated in the medial wall of the hemisphere between the anteriorly located areas denominated 10 and 24, and the posteriorly located rostral tip of the corpus callosum. This area 25 has a definite layer IV, contrary to minipig and human. In contrast, macaque’s area 24 is agranular and increasingly simpler in structure when approaching the corpus callosum. Moreover, the neurons in Walker´s area 24 are medium-sized and irregularly arranged. It seems, thus possible, that Walker’s area 25 resembles more well-differentiated granular cortical areas similar to human mPFC, whereas a less organized area more likely to correspond to the human area 25 is included in Walker’s area 24. This would be in concordance with the elaborate work of Carmichael and Price (Carmichael and Price 1994) on the macaque monkey prefrontal cortex using six different staining protocols. They also found an agranular area 25 caudal to a well-defined granular cortex, termed area 10 on the medial hemisphere. Their area 25 lacks clear radial orientation and has a prominent layer V and less dense layer II/III with the inner border of the cortex concluded with a thin layer VI. They give no description of an even less differentiated area caudal to area 25, which could correspond to area 33; however, their illustrations indicate an area termed tenia tectum, which classically is a poorly differentiated cortex although more ventrally located. It remains unelucidated whether this area corresponds to one or both of our areas nominated area 33 and IG, respectively. In the Macaca mulatta), area 25 is similarly located as in the Macaca fascicularis though, it seems as it extends ventrally into the orbital prefrontal region (Vogt et al. 1987; Barbas and Pandya 1989). The minipig area 25 abuts ventrally the olfactory tubercle and anteriorly to a thicker and more differentiated complex cortex, which we termed the mPFC (Bjarkam et al. 2017a). General features with gradually increasing laminar differentiation and cortical thickness when moving away from the septum and corpus callosum remain applicable in the minipig although the non-human primate area 25 in contrast to our findings seems to have more compact and denser infragranular layers V and VI.
The cytoarchitecture of the swine telencephalon has previously been investigated by Stephan (1951) who unfortunately did not include a separate description of the area termed 25, although his illustrations of the medial surface of the wild boar hemisphere depict such an area ventral to the rostral part of corpus callosum (Stephan 1951). This area is rather large and situated anterior to what is believed to be the septal area. Further rostral, this area abuts an even larger region marked area 8, which is considered granular cortex covering the bulk of the medial frontal surface (Stephan 1951). The cytoarchitecture of the Göttingen minipig prefrontal cortex has likewise been examined previously (Jelsing et al. 2006). In this study, an area denoted infralimbic cortex is recognized ventral to the rostral tip of the corpus callosum. This infralimbic area is not extensively described but does have clusters of cells in layer II as well as vertical cell poor strips. Furthermore, Jelsing et al. (2006), identified a periallocortical area and an IG dorsal to the corpus callosum in accordance with our findings on BA33 and IG. As in our studies (see also Bjarkam et al. 2017a), Jelsing et al. (2006) found the whole of the medial prefrontal area to be agranular, unlike Stephan (1951).
Area 25 is conventionally allocated to the cingulate cortex due to its pregenual localization and agranular appearance and its connectivity with the amygdala (Vogt et al. 2004). Jelsing has likewise included his infralimbic area in the anterior cingulate. We include the sgC in the pericallosal lobe of the Göttingen minipig, which also encompasses cytoarchitectonic distinct anterior cingulate (agranular cortex), posterior cingulate (dysgranular cortex), and retrosplenial cortex (real granular cortex) (Bjarkam et al. 2017a).
Neuronal Tract Tracing
The small size of the minipig sgC and the anatomical location close to the midsagittal level just below the rather large superior sagittal sinus made stereotaxic targeting difficult. Accordingly, only 3/8 animals had an injection in the sgC (Fig. 6), and some leakage to the ventricular system was subsequently observed in all traced animals.
It was furthermore evident when examining the sections with the fluorescence microscope that the retrograde tracing resulted in nicely labelled neuronal somata and some nerve fibers (Fig. 7), whereas the anterograde labelled terminals were minute and hard to distinguish from background staining (Fig. 8). Hence the anterograde tracing had to be interpreted on BDA-developed sections. The problems regarding the anterograde tracing could be circumvented by the performance of retrograde tracing from areas and structures believed to relate to the area 25. However, the limitations of this time-consuming and resource-demanding method are obvious. By selecting the areas to trace from, one has already opted out some of the possible connections and excluded “unexpected” results. Furthermore, such a procedure would require a large number of animals, and we had already tried to limit the number of used animals by combining the antero- and the posterograde tracing procedure for ethical and economic reasons.
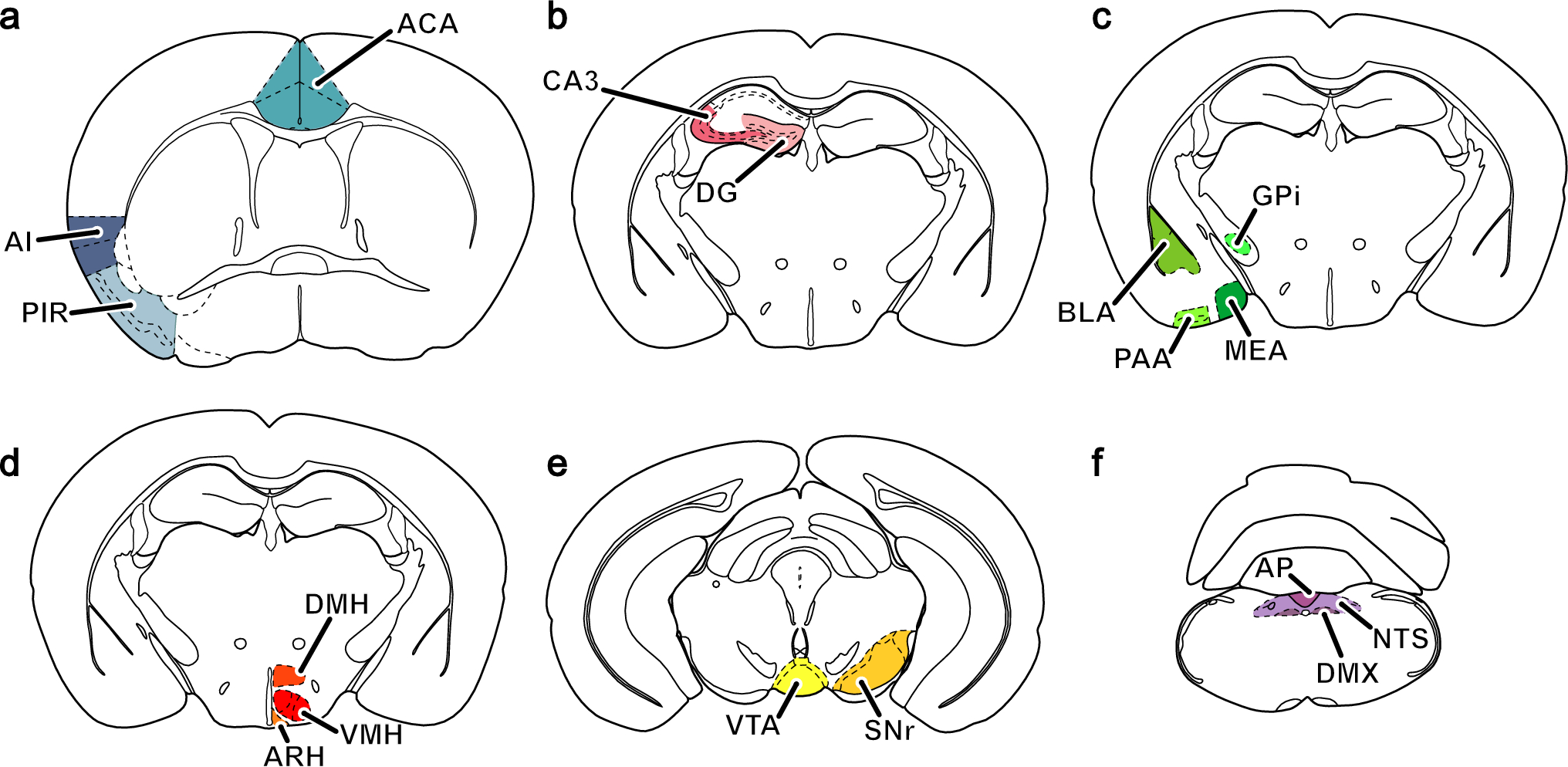
Despite the methodological problems outlined above, we found prominent retrograde connections from several cortical areas. This tracing was mainly confined to the neighbouring ventral and medial prefrontal areas, whereas more sparse connectivity was seen to the cingulate, insular, and perirhinal cortices. Strong projections to the hippocampal regions were observed as well as moderate to weak labelling in the amygdala.
Retrograde connectivity was likewise evident in the anterior, medial and dorsal aspects of the thalamus and posterior hypothalamus. Further caudally afferent connections with the ventral and dorsolateral periaqueductal grey, the ventral tegmental area and the substantia nigra appeared as well as some labelling in the midline raphe nuclei at mesencephalic and pontine levels.
Efferent connectivity included adjacent cortical areas in the cingulate and medial prefrontal cortices, the hippocampus, the caudate nucleus and less prominent tracing to the thalamus and hypothalamus.
Comparison with other tracing studies
Several studies have examined the connectivity of area 25 and the similar rat infralimbic cortex demonstrating a rich and diverse connectivity pattern (Vogt and Pandya 1987; Barbas and Pandya 1989; Takagishi and Chiba 1991; Condé et al. 1995; An et al. 1998; Ongür et al. 1998; Freedman et al. 2000; Chiba et al. 2001; Vertes 2004; Vogt and Vogt 2004). From these studies, it is clear that area 25 has widespread connections within the frontal cortex, especially the orbitofrontal cortex, as described in our study and reported in different non-human primates and rats (Vogt and Pandya 1987; Barbas and Pandya 1989; Condé et al. 1995). Anterograde tracing with tritiated amino acids injected in different orbital cortices of the rhesus monkey resulted in tracing to area 25 as well as other medial prefrontal cortical areas and adjacent orbitofrontal areas. Injections located in more dorsolateral frontal cortices (areas 46 and 8) did, however, not label terminals in area 25, which is in accordance with the absence of retrograde tracing to dorsolateral frontal cortices in our study (Barbas and Pandya 1989).
As seen in non-human primates and rats (Vogt et al. 1987; Vogt and Pandya 1987; Takagishi and Chiba 1991; Vertes 2004), we found reciprocal connections with the cingulate cortex. Similar to the findings in rat, we found some retrograde labelling in the insular cortex (Condé et al. 1995). This has also been reported in the rhesus monkey (Joyce and Barbas 2018, 2023); furthermore Vogt et al. found that the CgA area 24 in the rhesus monkey received afferents from the insular cortex (Vogt and Pandya 1987).
In the temporal lobe, we found retrograde tracing to the medial aspects of the amygdala. Projections from the amygdala have also been documented in the rhesus monkey and rat primarily located to the basolateral and basomedial nuclei (Vogt and Pandya 1987; Barbas and De Olmos 1990; Condé et al. 1995). We are at present not able to clarify whether these connections in the minipig is reciprocal, however, efferent projections from area 25 to the amygdala is well described in the primate and rat (Hurley et al. 1991; Takagishi and Chiba 1991; Freedman et al. 2000; Chiba et al. 2001; Vertes 2004). Finally, we observed reciprocal connections to the hippocampus and retrograde labelling to the subiculum and the entorhinal cortex, which also is reported in non-human primates (Vogt et al. 1987; Condé et al. 1995). However, hippocampus is not reported as a projection area of area 25 in the Japanese monkey, Macaca fuscata (Chiba et al. 2001) or the rat infralimbic area (Hurley et al. 1991). This constitutes a major difference between our study and the latter two studies. One feasible explanation is that our tracer injections are not only confined to the minipig sgC and hence the observed anterograde labelling in the hippocampus could arise due to tracing from adjacent septal areas.
Thalamic projections to area 25 in non-human primates arise primarily in the mediodorsal part of thalamus and midline nuclei and less from more anterior parts of the thalamus (Vogt et al. 1987). Similar findings are seen in the rat, but projections from the anteromedial nucleus in thalamus are also reported (Condé et al. 1995). We found consistent with these results retrogradely labelled neurons in the mediodorsal and anterior aspects of the minipig thalamus. Anterograde tracing from area 25 in the minipig yielded denser labelling in dorsal and midline nuclei and weaker labelling in the mediodorsal aspects. Efferent projections from area 25 in non-human primates and rats to thalamus are found in the paraventricular part of thalamus and the mediodorsal part (Takagishi and Chiba 1991; Freedman et al. 2000; Chiba et al. 2001; Vertes 2004).
Reciprocal connectivity between area 25 and hypothalamus is widespread in the macaque monkey as found by Ongür et al. (Ongür et al. 1998), whereas in the minipig, we found a more restricted retrograde labelling, confined to dorsal and posterior parts of the hypothalamus. Our results are more consistent with data on the rat infralimbic area, in which the hypothalamic anterograde projections are more limited. However, in the rat the projections arise primarily in the lateral hypothalamic area (Conde et al. 1995). The projections from area 25 terminate throughout the macaque hypothalamus, but most densely in the ventromedial nucleus (Ongür et al. 1998; Chiba et al. 2001), whereas in the rat, efferent projections terminate in more lateral and dorsomedial aspects of the hypothalamus. Unfortunately, our anterograde results were technically not conclusive about the efferent projections from the minipig area 25 to hypothalamus.
Anterograde tracing to the caudate nucleus is recognized in our study and is consistent with findings in the studies conducted in both non-human primates (Freedman et al. 2000), where tracing is found in the ventromedial caudate nucleus and dorsomedial caudate, and in rats, where the infralimbic area shows projections to the medial caudate and putamen (Vertes 2004).
Further caudal projections from area 25 in non-human primates are found in different structures in the brainstem, such as the ventral tegmental area, dorsolateral periaqueductal grey, dorsal raphe nuclei and locus coeruleus. Similar projections from the infralimbic area in the rat are found but, in addition, projections continue caudally to the nucleus ambiguous and nucleus of the solitary tract (Hurley et al. 1991; Takagishi and Chiba 1991). Such direct autonomic projections from area 25 in the non-human primates have not been reported (Freedman et al. 2000; Chiba et al. 2001) and thus we would not expect to find such connections in the minipig. However, our study did not provide any results regarding the efferent projections to the minipig’s brainstem; therefore, this cannot be either dismissed or verified. Different areas in the brainstem of the minipig does seem to project to the area 25 such as the ventral tegmental area, the periaqueductal grey, the rostral raphe nuclei and substantia nigra, which is consistent with findings in the rat (Condé et al. 1995) this.
Functional considerations
Traditionally reciprocal connectivity with the anterior and mediodorsal nucleus of the thalamus has been used to define prefrontal cortical areas and the anterior cingulated cortex, respectively (Devinsky et al. 1995; Vogt et al. 2004; Zilles 2004). In humans the anterior part of the cingulate cortex is classified into the perigenual anterior cingulate cortex (pACC), which comprises the cytoarchitectonic areas termed 32, 24, 25 and the rostral part of area 33 (Vogt et al. 2004). Functionally, the pACC is further subdivided into an area located ventral of the rostrum of the corpus callosum, and the subgenual subregion lying more rostral and ventral to genu corporis callosi comprising the cytoarchitectonic areas 33, 25, 24a and the caudal extreme of area 32 (Vogt et al. 2004).
Comparison of the minipig area 25 to the infralimbic cortex in rats shows some general similarities, but also clear differences (Vogt and Vogt 2004). They both have poor laminar differentiation and thin cortical thickness. Furthermore, the rat layer III is poorly differentiated whereas layer V in the rat is uniform and thick. In the minipig, the discrimination between layer III and V is less obvious, as both layers generally contains evenly sized and dispersed cells (Fig. 5). Importantly although this article does not include behaviour; in rodents, infralimbic cortex is associated with extinction and inhibition of fear responses, whereas in primates, area 25 is not and in fact promotes amygdala activity related to fear and anxiety. (Roberts and Clarke 2019; Wallis et al. 2017).
Our tracing data indicates reciprocal connections between the medial, dorsal and anterior aspects of the thalamus and the subgenual cortex in the minipig, which further strengthens the view that the minipig possesses a cortical area in its subgenual region, homologous to the human area 25. It can furthermore not be disregarded that we found projections from the amygdaloid complex to area 25 in the minipig, which also is found in non-human primates and rats and hence believed to correspond to human connectivity (Vogt et al. 1987, 2004; Vogt and Pandya 1987). Accordingly, it is conceivable that the Göttingen minipig has a homologous subgenual region, which may have similar functional properties. Based on connectional data showing efferent connections from the infralimbic area in the rat to autonomic brainstem nuclei and electrical stimulation of the subgenual subregion eliciting autonomic responses in humans, this region is thought of as a visceromotor control region (Vogt et al. 2004). Considering these data, it is interesting to speculate if this in fact represents the anatomical framework responsible for the autonomic deficits seen in severely depressed individuals.
Interestingly, we also found some rather prominent retrogradely labelled neurons in the ventral tegmental area and less intensively labelling in the pontine and mesencephalic raphe nuclei, which also have been reported in the rat. It is of particular interest that one of the most widely propagated theories concerning the pathophysiology and pharmacological treatment of the major depressive disorder is centred around disturbances in monoaminergic pathways known to originate in part from the identified brain stem structures (Nestler et al. 2002a, b).




留言 (0)