
記住我









Sham control (Ctrl) or BCAS surgeries were performed in C57BL/6 J male mice (Fig. 1A, B) and BCAS-induced cerebral hypoperfusion was assessed by PeriCam laser speckle contrast analysis. We measured the rCBF in the cerebral hemisphere including cerebral frontal cortex, and cerebellum regions (Fig. 1C, D, E, F). Figure 1C illustrates representative PeriCam laser speckle images of rCBF before, 10 min, 3 days, 7 days, 14 days, and 28 days after either Sham or BCAS surgery. As shown in Fig. 1C, D, there were no significant changes in rCBF in in the cerebral hemispheres of the Sham Ctrl group between the pre-surgery baseline and 3–28 days post-surgery time points (p > 0.05, Fig. 1C, D). In contrast, the BCAS group exhibited an approximately 20% decrease in hemispheric rCBF at 10 min post-BCAS surgery (**p < 0.01) and this cerebral hypoperfusion sustained at 3–28 days post-BCAS (*p < 0.05, **p < 0.01, respectively, Fig. 1C, D). In addition, we have analyzed CBF changes in the cerebral frontal cortex and cerebellum in the Sham and BCAS brains (Fig. 1E, F and Supplemental Figure S1C, D). It shows that BCAS male mice displayed reduced rCBF in the frontal cortex regions approximately by 25% (10 min post-BCAS) and by 18% (3 days post-BCAS) (****p < 0.0001, *p < 0.05, Fig. 1E). Additionally, the rCBF in the cerebellum also decreased significantly at 10 min (by ~ 29%) and at 3 days (by 21%) post-BCAS (****p < 0.0001, *p < 0.05, Fig. 1F). By 7 days post-BCAS, the rCBF in both regions had recovered to > 90% of the baseline levels (p > 0.05, Fig. 1E, F).
Since vascular dementia appears to be more common in males in clinical settings, we assessed whether male and female mice displayed different responses after BCAS. In female mice, the rCBF in the cerebral frontal and cerebellum regions decreased by approximately 24% and 20%, respectively, at 10 min post-BCAS surgery (*p < 0.05, **p < 0.01, Figure S1C, D). By 3 days post-surgery, the rCBF in both regions had recovered to > 90% of the baseline levels (p > 0.05, Figure S1C, D). The female mice dropped approximately 4–5% more in the CBF of hemispheres than in males at 10 min post-BCAS (Supplemental Figure S1E, p > 0.05) but followed with a faster recovery (a 9%-10% CBF increase) during the compensation period from 3 to 28 days post-BCAS (Figure S1 E, p > 0.05). This sex difference is consistent with human studies, as vascular dementia, which is caused by a reduction of blood flow to the brain, is more frequent in males [27], perhaps pointing to reduced compensation of blood flow after neurovascular events in males. Due to early recovery of the rCBF in female mice post-BCAS, we only conducted experiments in male mice in the rest of the study.
BCAS induces changes of ChP SPAK-NKCC1 protein complexWe then investigated whether BCAS-induced any changes of ChP structures. Figure 2A illustrate three regions of interest (ROI 1–3) selected from each side of the LVCP. We examined whether BCAS triggers changes of the ChP secretory ion transport protein complex SPAK-NKCC1. Sham ChP show total non-phosphorylated SPAK and NKCC1 protein enriched at the apical (luminal) membrane of ChP epithelial cells, but less in the basolateral membrane. In contrast, BCAS mice displayed sustained upregulation of both total NKCC1 protein (~ 1.85-, ~ 2.55- or 2.83-fold, respectively) and pNKCC1 protein (~ 1.88-, ~ 1.85-, ~ 2.65-) at 4-, 6- and 8-weeks post-BCAS (*p < 0.05, **p < 0.01, Fig. 2B, C, D, E.). However, the regulatory serine-threonine kinases SPAK and phosphorylated SPAK (pSPAK) showed opposite responses, with initial significant reduction of apical SPAK expression (by 46%) (**p < 0.01, Fig. 2B, C), but elevated pSPAK protein expression by twofold at 4-weeks post-BCAS (*p < 0.05, Fig. 2D, E). At 6-weeks post-BCAS, SPAK levels significantly increased to twice that of the Sham Ctrl, and by 8-weeks post-BCAS, SPAK immunostaining intensity nearly returned to the basal levels (**p < 0.01, *** < 0.001, Fig. 2B, C). pSPAK began to return to the basal levels starting at 6-weeks post-BCAS (*p < 0.05, Fig. 2D, E). Taken together, these data demonstrate that BCAS causes dynamic changes of ChP secretion function-related proteins SPAK and NKCC1 at 4–8 weeks post-BCAS.
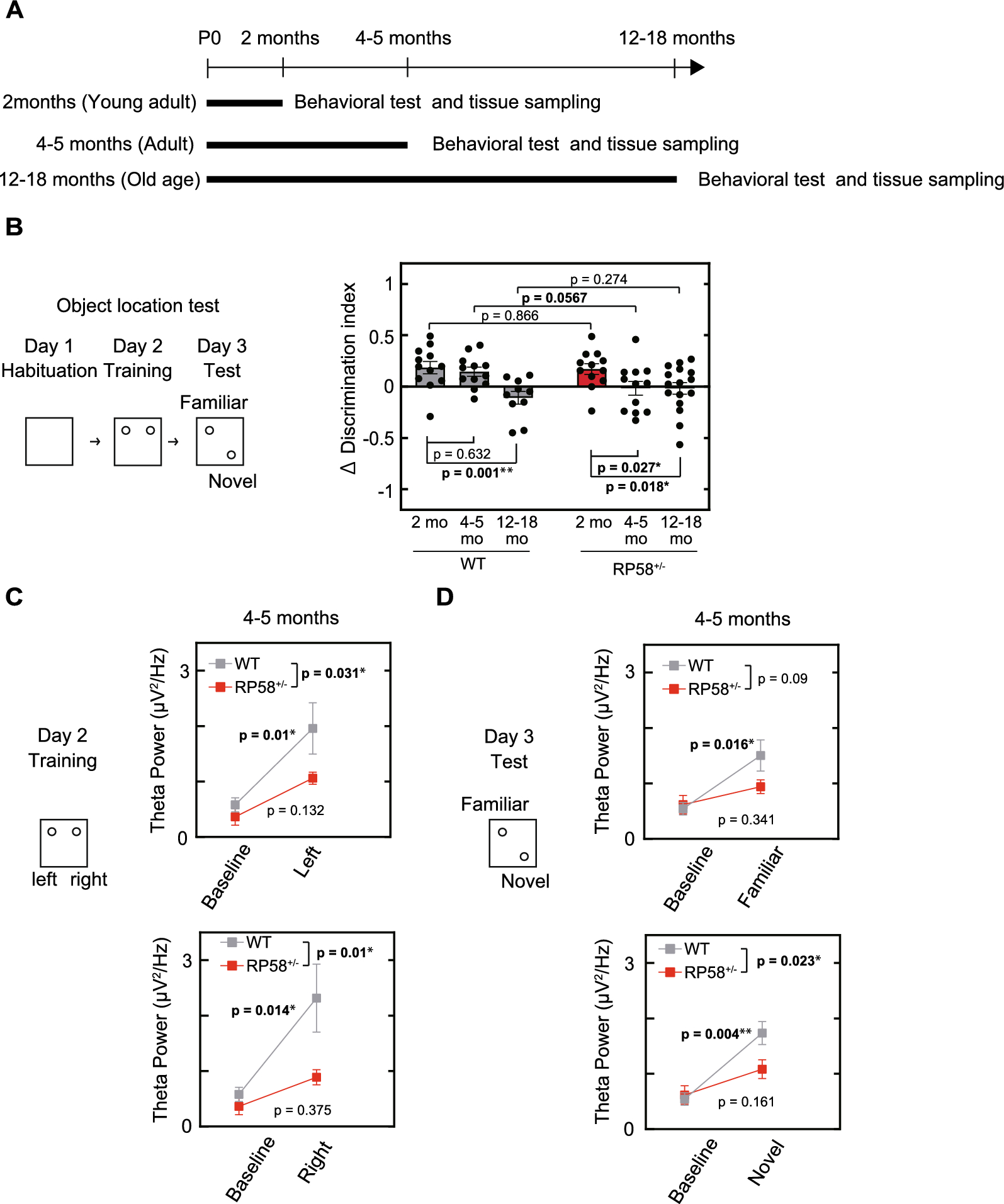
BCAS mice display neurological behavioral deficit and sensitivity to SPAK inhibitor ZT-1a treatment at 2–4 weeks post-BCASWe next assessed whether BCAS triggered neurological deficits in mice, and if they were sensitive to SPAK inhibitor ZT-1a treatment. C57BL/6 J male mice were subjected to sham or BCAS surgery with treatment regimens with either BCAS + Veh (100% DMSO, 2 ml/kg body weight/day) or BCAS + ZT-1a (5 mg/kg body weigh/day, i.p.) at 14–28 days post-surgery (Fig. 3A). The neurological function was determined with Open Field (OF) and Y maze tests. In the OF tests, the total distance traveled is used to assess the exploratory behavior and activity inclination of mice, while the number of vertical activity counts is used to evaluate exploratory behavior and curiosity [29, 30]. Compared to the sham group, the BCAS + Veh group showed a marked reduction in both total travel distance (by 58%, **p < 0.01) and vertical activities (by 54%, *p < 0.05, Fig. 3B, C), revealing reduced locomotor activity and exploratory drive at 4-weeks post-BCAS. However, the BCAS + ZT-1a group displayed similar performance on total travel distance and vertical activity counts as the sham group (Fig. 3B, C). These findings suggest that BCAS-induced anxiety and/or motor dysfunction can be effectively mitigated by pharmacological blockade of SPAK protein using the SPAK inhibitor ZT-1a at 2–4 weeks post-BCAS. In testing changes of spatial working memory in these mice in the Y-maze test, there were no differences in the total arm entries or spatial working memory among the three groups of mice (p > 0.05, Fig. 3D).
Fig. 3
Administration of SPAK inhibitor ZT-1a at 2–4 weeks post-BCAS attenuates neurological behavioral deficits in mice. A Experimental protocol. C57BL/6 J mice underwent sham or BCAS surgery. BCAS mice received either vehicle (100% DMSO, 2 ml/kg body weight/ day) or ZT-1a (5 mg/kg body weigh/day) via intraperitoneal injection (i.p.) from 14 to 28 days. Arrowheads: indicate the time points at which drug treatment, behavior, IF, WB or flow cytometry were performed. B Illustration of animal total distance traveled in the Open field test. C Summary of travel distance and vertical movement counts. Data are represented as mean ± SD. n = 7–8 mice. *p < 0.05, **p < 0.01. One-way analysis of variance (ANOVA). D Y-maze spontaneous alternation test. The percentage of spontaneous alternation and total entry in Y maze test was shown. Data are mean ± SD, n = 6–8 mice. *p < 0.05. One-way analysis of variance (ANOVA)
Considering that cerebral hypoperfusion has been reported to induce optic nerve damage and visual impairment [31], which could affect the outcomes of neurological behavioral tests, we assessed reactive astrocyte activation in optic nerves of the Sham and BCAS-Veh, and BCAS-ZT-1a brains. As shown in Supplemental Figure S3, the optic nerve of Sham mice exhibited low astrocytic GFAP mean fluorescence intensity (MFI). Some localized elevation in astrocytic GFAP MFI was detected in the optic nerves of some BCAS-Veh mice, however, overall differences between the Sham and BCAS-Veh mice are not statistically significant (p > 0.05, Figure S3A, B). These data suggest that the BCAS model in mice did not cause severe optic nerve damage at 4 weeks post-BCAS, which is consistent with previous report on a mild demyelination in the optic tract post-BCAS [32]. The lack of the optical nerve damage in the BCAS mouse model could be due to preserved blood flow in the ophthalmic branch artery [33, 34]. Interestingly, the optic nerves of BCAS-ZT-1a mice displayed significantly attenuated GFAP MPI, compared to the Sham or BCAS-Veh group while without performance differences among the three groups in the Y-maze test. These data further imply that potential contribution of the differences in optic nerve damage or visual capacity is negligible in assessing the OFT and Y-maze tests at 4-weeks post-BCAS.
SPAK inhibitor ZT-1a reduces stimulation of ChP pSPAK-pNKCC1 complex and pNF-kB protein at 2–4 weeks post-BCASWe then evaluated the effect of pharmacological inhibition of SPAK protein on SPAK-NKCC1 complex activation and NF-κB activation. In the Veh-treated BCAS brain, the expression levels of pSPAK and pNKCC1 (arrows) in the ChP were elevated by twofold and threefold, respectively, compared to the Sham Ctrl (*p < 0.05 for pSPAK, ***p < 0.001 for pNKCC1, Fig. 4A, B). In contrast, treatment with ZT-1a prevented such upregulation of pSPAK and pNKCC1 in the ChP epithelial cells of the BCAS + ZT-1a brains, which showed at 80% and 70% of the Sham Ctrl levels (Fig. 4A, B). This is significantly different from data in the BCAS + Veh ChP (**p < 0.01 for pSPAK, ****p < 0.0001 for pNKCC1). SPAK functioning as an inflammatory node involves NF-κB activation [12, 13]. We further evaluated changes of pNF-κB protein in ChPs from the BCAS + Veh and the BCAS + ZT-1a brains. NF-κB dimers (p65/p50) are normally sequestered in the cytoplasm by inhibitory factor-KB proteins (IκBα/β/ε). However, when the IκB kinase (IκK) becomes activated, it phosphorylates IκB, causing its dissociation. This enables the NF-κB complex to translocate from the cytosol to the nucleus, initiating transcription [18]. Sham Ctrl showed low expression levels of NF-κB or pNF-κB in the choroid plexus epithelial cells (Fig. 4C, D). But, in the Veh-treated BCAS brain, pNF-κB (Ser536) in the choroid plexus epithelial cells was increased significantly (2.29-fold), compared to the Sham Ctrl (****p < 0.0001, Fig. 4D), while the rise in NF-κB expression was less pronounced (p > 0.05). Administration of the SPAK inhibitor ZT-1a post-BCAS reduced the upregulation of pNF-κB by 56% (****p < 0.0001, Fig. 4D). To further detect changes of nuclear p-NF-kB signals, using Imaris reconstruct tool (the spot tool), we quantified changes of p-NF-κB immunostaining signals within the DAPI+-reconstructed structures (Fig. 4D). For each stack image, the “total number of p-NF-κB immunostaining spots” were divided by the “total number of DAPI+ surfaces” to represent p-NF-κB puncta density in the nucleus. Figure 4D showed that the BCAS + Veh ChP epithelial cells showed a 1.39-fold increase in the p-NF-κB puncta density compared to the Sham Ctrl (*p < 0.05, Fig. 4D). But the BCAS + ZT-1a ChP illustrated a 35% reduction of the p-NF-κB puncta density compared with the BCAS + Veh ChP (p = 0.1681, Fig. 4D). These data demonstrate the involvement of SPAK in the post-BCAS induced activation of pNF-κB inflammatory pathway.
Fig. 4
ZT-1a treatment at 2–4 weeks post-BCAS attenuates SPAK-NKCC1 complex and pNF-kB activation. A Representative immunofluorescent staining image of pSPAK and pNKCC1 in the Sham or BCAS + Veh, BCAS + ZT-1a LVCPs. Arrowheads: low level expression of pSPAK or pNKCC1. Arrows: elevated pSPAK or pNKCC1 expression. B Quantification summary. Data are mean ± SD (n = 4 mice). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. One-way analysis of variance (ANOVA). C. Representative immunofluorescent staining Max IP image of NF-κB and pNF-κB, and 3D cell reconstruction images of nuclear pNF-kB. D Quantification summary. Data are mean ± SD (n = 4–6 mice). *p < 0.05, ****p < 0.0001 vs Sham. One-way analysis of variance (ANOVA), One Sample t-test or unpaired t-test
Changes of the SPAK-NKCC1 complex in LVCP at 4 weeks post-BCAS were also analyzed in immunoblots. LVCP homogenates from the Sham, BCAS + Veh and BCAS + ZT-1a brains showed similar levels of expression of SPAK and pSPAK proteins, NKCC1 and pNKCC1 proteins or claudin-1 (p > 0.05, Supplemental Figure S4A, B, C, D). With LVCP tissue homogenates (no isolation of cytosolic and nuclear fractions due to small cell numbers), no differences in NF-kB expression were detected among Sham, the BCAS + Veh and the BCAS + ZT-1a ChPs (p > 0.05, Figure S4E, F). The discrepancies between findings of immunofluorescence staining and immunoblotting approaches could be that Western blotting probes protein levels (SPAK, NKCC1, NF-kB, pNF-κB) across the entire LVCP, while immunofluorescence analysis enables to detect localized changes within specific ChP ROI regions of epithelial cells. The discrepancy between the immunostaining and immunoblot data could also result from heterogenous CBF hypoperfusion within the LVCP, the heterogenous damage of ChP can be detected by the immunostaining analysis of specific ROI but got lost in the whole LVCP homogenate preparation.
Pharmacological inhibition of the SPAK–NKCC1 complex at 2–4 weeks post-BCAS reduces brain immune cell infiltrationWe further investigated whether BCAS leads to neuroinflammation at 14-28d post-surgery and whether pharmacological inhibition of SPAK attenuates such events. Changes of immune cell infiltration in the cerebral hemisphere or the ChP in the BCAS + Veh or BCAS + ZT-1a mice were quantified at day 28 post-surgery via flow cytometry analysis. Compared to the Sham Ctrl, the BCAS + Veh cerebral hemisphere exhibited a significant increase in the number of CD11b+CD45+hi myeloid cells (by twofold, *p < 0.05, Fig. 5A). In contrast, 50% reduction was detected in the number of CD11b+CD45+lo myeloid cells (**p < 0.01, Fig. 5A). Additionally, a 1.63-fold increase in CD11b+CD45+/Ly6G+ neutrophils (*p < 0.05, Fig. 5C) and a 35% decrease in CD11b+CD45+lo/CD206+ anti-inflammatory myeloid cells was detected in the BCAS + Veh cerebral tissues (*p < 0.05, Fig. 5B). No significant changes in CD3+ T cells were observed (p = 0.6230, Fig. 5C), compared to the sham Ctrl. In contrast, the BCAS + ZT-1a cerebral hemisphere tissues displayed reduced quantity of CD11b+CD45+hi cells (p = 0.0549, Fig. 5A), increased anti-inflammatory CD11b+CD45+lo/CD206+ myeloid cells (p = 0.1750, Fig. 5B), and reduced infiltration of neutrophils (**p < 0.01, Fig. 5C).
Fig. 5
Administration of ZT-1a at 2–4 weeks post-BCAS reduces immune cell infiltration into hemisphere and ChP. A Representative flow cytometric plots of CD11b+CD45+hi or CD11b+CD45+lo myeloid cells from cerebral hemisphere at 4 weeks post-BCAS surgery with the percentage of total number of CD11b+CD45+hi or CD11b+CD45+lo myeloid cells in the single cells. Data are mean ± SD (n = 7 mice). *P < 0.05, **p < 0.01. One Sample t-test. Unpaired t-test. B Percentage of anti-inflammation CD11b+CD45+hi/CD206+ and CD11b+CD45+lo/CD206+ cells. Data are mean ± SD (n = 7 mice). *p < 0.05. One Sample t-test. Unpaired t-test. C. The percentage of total number of CD3+ lymphocyte in the single cells and CD11b+CD45+/Ly6G+ neutrophils. Data are mean ± SD (n = 8 mice). *p < 0.05, **p < 0.01. One Sample t-test. Unpaired t-test. D Representative flow cytometric plots of CD11b+CD45+hi or CD11b+CD45+lo myeloid cells from the LVCP and the percentage in the single cells. Data are mean ± SD (n = 8–9 mice). One Sample t-test. Unpaired t-test. E Percentage of anti-inflammatory CD11b+CD45+hi/CD206+ and CD11b+CD45+lo/CD206+ cells. Data are mean ± SD (n = 8–9 mice), *p < 0.05, ** < 0.01. One Sample t-test. Unpaired t-test. F The percentage of total number of CD3+ lymphocyte and CD11b+CD45+/Ly6G+ neutrophils in the single cells. Data are mean ± SD (n = 8–9 mice), *p < 0.05. One Sample t-test. Unpaired t-test
In flow cytometry analysis of immune cell infiltration in the ChPs, the landscape is very different. Compared to the sham Ctrl, the BCAS + Veh ChPs exhibited less increase in CD11b+CD45+hi and CD11b+CD45+lo myeloid cells (p = 0.1977, p = 0.1259, Fig. 5D). However, a significant reduction of CD11b + CD45+hi/CD206+ (26%) and CD11b+CD45+lo/CD206+ (18%) myeloid cells were detected (*p < 0.05, Fig. 5E). An obvious increase in CD11b+CD45+/Ly6G+ neutrophils (*p < 0.05, Fig. 5F) and a 1.68-fold increase in CD3+ T cells (*p < 0.05, Fig. 5F) were detected in the BCAS + Veh ChPs at 4-weeks post-BCAS. Treatment with ZT-1a normalized the level of anti-inflammatory CD11b+CD45+hi/CD206+ and CD11b+CD45+lo/CD206+ myeloid cells near to the sham Ctrl levels (p = 0.1198, **p < 0.01, Fig. 5E), while also reducing the infiltration of neutrophils and CD3+ T cells into the ChP (p = 0.0797, p = 0.1489, Fig. 5F).
To further assess changes immune cell infiltration in the cortex, peri-ventricle, and ChP regions, we performed immunofluorescence staining for Iba1+ cells in these regions. Compared to the Sham group, the number of Iba1+ cells was significantly increased in the peri-ventricle region of the BCAS + Veh brains (*p < 0.05, Supplemental Figure S5A, C), a less profound elevation was detected in the cortex and ChP (p > 0.05, Figure S5A, C). Consistently, the BCAS + ZT-1a brains (the cortex, peri-ventricle, or ChP regions) did not show statistically significant changes in Iba1+ counts, compared to the Sham Ctrl (p > 0.05, Figure S5A, C). In summary, these findings clearly demonstrate a role of the SPAK in immune cell infiltration in BCAS cerebral tissues and ChP.
ZT-1a administered at 4–8 weeks post-BCAS remains protectiveWe further investigated whether administration of SPAK inhibitor ZT-1a during 4–8 weeks post-BCAS remains protective against hypoperfusion-induced damage. The BCAS + Veh mice (100% DMSO, 2 ml/kg body weight/day) or BCAS + ZT-1a mice (5 mg/kg body weight/day, i.p.) received treatment regimens from day 28 to 56 post-BCAS, as shown in Fig. 6A. The sham Ctrl group did not show significant changes in rCBF between the pre-surgery baseline throughout 8-weeks post-surgery (p > 0.05). In contrast, the BCAS + Veh mice exhibited an approximate 40% reduction in rCBF at 8-weeks post-BCAS (***p < 0.001, Fig. 6B, C). Interestingly, in the BCAS + ZT-1a mice, the rCBF was preserved at 83% of the pre-surgery level, showing significantly less hypoperfusion (*p < 0.05, Fig. 6B, C). In assessing their performance in the Y-maze for the Spontaneous Alternation and Novel Spatial Recognition (NSR) tests, the BCAS + Veh mice showed 20% reduction in the spontaneous alternation test, indicating deficits in spatial working memory (*p < 0.05, Fig. 6D), which was absent in the BCAS + ZT-1a group (**p < 0.01, Fig. 6D). These findings were further supported by the Recognition Index data of the NSR test, which was defined as the duration in novel arm divided by the total duration spent on both familiar and novel arms, to represent changes in the mice’s curiosity towards previously unvisited areas. The Sham Ctrl group exhibited an intact working memory with a higher recognition index. In contrast, the BCAS + Veh group showed 34% reduction in the Recognition Index (**p < 0.01, Fig. 6E). The BCAS + ZT-1a mice did not display any differences from the Sham Ctrl group (Fig. 6E). Taken together, these findings clearly show that pharmacological blockade of SPAK protein with ZT-1a at 4–8 weeks post-BCAS can effectively improve the recovery of CBF as well as preserving spatial working memory.
Fig. 6
Protective effects of SPAK inhibitor ZT-1a at 4–8 weeks post-BCAS. A Experimental protocol. C57BL/6 J mice underwent sham, BCAS + Veh (100% DMSO, 2 ml/kg body weight/ day, i.p.), or BCAS + ZT-1a (5 mg/kg/day, via intraperitoneal injection, i.p.) at 28-56d. Arrowheads: indicate the time points at which drug treatment, CBF or behavioral test were performed B Representative periCam PSI images of rCBF at 56 days (8-weeks) after surgery. Dashed circle: region of interest (ROI). C Quantification analysis of rCBF. Data are expressed as mean ± SD. n = 4 mice. *p < 0.05, ***p < 0.001 vs. Sham. One-way analysis of variance (ANOVA). D The percentage of spontaneous alternation and total entry in Y maze test. n = 10–12 mice. *p < 0.05, **p < 0.01. One-way analysis of variance (ANOVA). E The recognition index of the Novel Spatial Recognition (NSR) test. Data are represented as mean ± SD. n = 8–10 mice. *p < 0.05, ** < 0.01 vs. sham. One-way analysis of variance (ANOVA)
ZT-1a-mediated sustained attenuation of immune cell infiltration into the cortical hemisphere and ChP treatmentWe then assessed whether post-BCAS ZT-1a administration reduced immune cell infiltration in the cerebral hemisphere and ChP at 8-weeks post-BCAS. The flow cytometry analysis revealed that at 8-weeks post-BCAS, there was a persistent increase in the number of CD11b+CD45+hi myeloid cells, but not the CD11b+CD45+lo myeloid cells, in the cerebral cortical hemisphere of the BCAS + Veh mice, compared to the sham Ctrl (**p < 0.01, Fig. 7A). Additionally, CD3+ T cells in the BCAS + Veh cerebral hemisphere remained significantly elevated (*p < 0.05, Fig. 7C), accompanied with a 25% decrease in CD11b + CD45+hi/CD206+ myeloid cells (*p < 0.05, Fig. 7B). In contrast, the immune cell infiltration profiles in the BCAS + ZT-1a cerebral hemisphere are significantly different, with a reduction of CD11b+CD45+hi (p < 0.05, Fig. 7A) and CD3+T cell infiltration (**p < 0.01, Fig. 7C) and an increase in anti-inflammatory CD11b+CD45+hi/CD206+ myeloid cells (p = 0.2835, Fig. 7B). Interestingly, in the ChP, the BCAS + Veh mice showed sustained 2.3-fold increase in CD11b+CD45+/Ly6G+ neutrophils, while no significant changes in anti-inflammatory myeloid cell or CD3+ T cell infiltration, indicating partial recovery from inflammatory responses by 8-weeks post-BCAS. In contrast, the CD11b+CD45+/Ly6G+ neutrophil infiltration into the ChP was absent in the BCAS + ZT-1a brains (**p < 0.01, Fig. 7F). These findings indicated the evolution of neuroinflammation between 4 and 8 weeks post-BCAS. Blocking the SPAK-NKCC1 complex with ZT-1a attenuated BCAS-induced immune cell infiltration into the cerebral hemisphere and ChP.
Fig. 7
ZT-1a treatment at 4–8 weeks post-BCAS attenuated chronic immune cell infiltration into the hemisphere and ChP. A Representative flow cytometric plots of CD11b + CD45 + hi or CD11b+CD45+lo myeloid cells in the single cells from hemisphere at 8-weeks post-surgery. Data are mean ± SD (n = 7 mice). *P < 0.05, **p < 0.01. One Sample t-test. Unpaired t-test. B Fold changes of anti-inflammation CD11b+CD45+hi/CD206+ and CD11b + CD45+lo/CD206+ cells. Data are mean ± SD (n = 7 mice). *p < 0.05. One Sample t-test. Unpaired t-test. C Fold changes of total number of CD3+ lymphocyte and CD11b+CD45+/Ly6G+ neutrophils. Data are mean ± SD (n = 7 mice). *p < 0.05, ** p < 0.01. One Sample t-test. Unpaired t-test. D Representative flow cytometric plots of CD11b+CD45+hi or CD11b + CD45+lo myeloid cells from the LVCP and the percentage in the single cells. Data are mean ± SD (n = 7–8 mice). One Sample t-test. Unpaired t-test. E Fold changes of anti-inflammatory CD11b+CD45+hi/CD206+ and CD11b+CD45+lo/CD206+ cells. Data are mean ± SD (n = 7–8 mice), p > 0.05. One Sample t-test. Unpaired t-test. F Fold changes of CD3+ lymphocyte and CD11b+CD45+/Ly6G+ neutrophils in the single cells. Data are mean ± SD (n = 7–8 mice), *p < 0.05. One Sample t-test. Unpaired t-test
Pharmacological inhibition of the SPAK–NKCC1 complex preserves the ChP blood-CSF barrier structure in BCAS brainsTo further investigate whether pharmacological inhibition of SPAK protein protects the blood-CSF barrier, we examined changes of claudin-1 expression in the ChP epithelial cells in the Veh vs ZT-1a treated mice at 4-weeks post-BCAS. In the Sham ChP, the normal location of claudin-1 TJ proteins was observed between the epithelial cells of the ChP (Fig. 8A). In the BCAS + Veh ChP, there was a significant increase in the immunofluorescence intensity of claudin-1, approximately 1.8 times higher than that of the Sham group (*p < 0.05, Fig. 8A, B), that especially appeared near the epithelial apical membranes (as indicated by sector-shaped box and arrow, *p < 0.05, Fig. 8A, B). IMARIS 3D reconstruction of these images illustrated that BCAS-induced loss of polarity of claudin-1 protein expression at the ChP epithelial cells (as indicated by arrowhead), with the increased volume of claudin-1+ stained structure than the Sham Ctrl (*p < 0.05, Fig. 8A, C), collectively indicating maldistribution of TJ proteins at the blood-CSF barrier after BCAS. In contrast, in the BCAS + ZT-1a ChPs, the immunofluorescence intensity of claudin-1 and its abnormal location at the apical membranes of ChP epithelial cells were decreased to a level like the Sham Ctrl (*p < 0.05, Fig. 8A, B). We further investigated changes in expression of claudin-5 and E-cadherin in the ChP at 4-weeks post-BCAS. In Sham ChP, the claudin-5 protein was primarily expressed in ChP epithelial cells (Fig. 9A). The immunofluorescence intensity of claudin-5 was significantly increased in the BCAS + Veh ChP, compared to the Sham group (by 2.7-fold, *p < 0.05, Fig. 9A, C). In contrast, such a change in claudin-5 in the BCAS + ZT-1a Chp is not significant (p = 0.0865, Fig. 9A, C). As for E-cadherin, it was abundantly distributed in the ChP epithelial cells (Fig. 9B), which was elevated in the BCAS + Veh group (p = 0.1407, Fig. 9B, D). Taken together, BCAS triggered changes of claudin-1, claudin-5, and E-cadherin proteins in a similar trend. To further assess if the permeability of the blood-CSF barrier is compromised at 4-weeks post BCAS, we conducted an initial evaluation by measuring albumin leakage into the CSF. We measured albumin levels in the CSF and serum from the Sham, BCAS + Veh, and BCAS + ZT-1a mice at 4 weeks post-BCAS. The ratio of CSF Albumin/Serum Albumin in the BCAS + Veh mice was 1.34-fold of the Sham mice (p = 0.4940, Fig. 9E). The BCAS + ZT-1a mice displayed a similar value in the CSF Albumin/Serum Albumin ratio (p = 0.3595). These data indicate that despite of BCAS-induced cellular structural changes in the blood-CSF barrier, its permeability for large proteins such as albumin (66.5 kDa) is not compromised (Fig. 10).
Fig. 8
Effects of ZT-1a on changes of ChP TJ protein expression after BCAS. A Representative immunofluorescent staining of claudin-1 in the sham, BCAS + Veh, or BCAS + ZT-1a brains (2–4 weeks post-BCAS) treatment in single-layer images, Z-Stacks Max IP, and 3D cell reconstruction of immunofluorescent staining images derived from these Z-stacks. Arrows and sector-shaped boxes: the ectopic localization of claudin-1 at the apical membrane of the ChP epithelial cells; Arrowheads: expression of claudin-1. B Quantification summary. Data are mean ± SD (n = 5 mice). *p < 0.05. One Sample t-test. Unpaired t-test. C Quantification summary. Data are mean ± SD (n = 5 mice). *p < 0.05. One Sample t-test. Unpaired t-test
Fig. 9
Effects of ZT-1a on changes of ChP claudin-5 and E-cadherin expression after 4 weeks post-BCAS. A Representative immunofluorescent staining images of claudin-5 in the sham, BCAS + Veh, BCAS + ZT-1a ChP (including single-layer images, Z-Stacks Max IP, and 3D cell reconstruction images derived from these Z-stacks). Rectangular boxes: indicate the regions presented in the Z-stacks maximum intensity projection (Max IP) and 3D reconstruction images; Arrowheads: indicate the expression of claudin-5. B Representative immunofluorescent staining images of E-cadherin. Rectangular boxes: indicate the regions presented in the Z-stacks maximum intensity projection (Max IP) and 3D reconstruction images. Arrowheads: indicate the expression of E-cadherin. C–E Quantification summary. Data are mean as ± SD (n = 5 mice). *p < 0.05. One Sample t-test. Unpaired t-test. Quantification of albumin in CSF and serum in E was by ELISA
Fig. 10
Schematic summary of the brain-ChP barrier damage after BCAS. The ChP, with tight junctions (TJ) at the apical membrane of the ChP epithelial cells, prevents inflammation by blocking immune cells from entering the brain. Na+-K+-Cl− cotransporter isoform 1 (NKCC1) and its regulatory serine-threonine kinase, SPAK, located at the apical membrane, are essential in regulating choroid plexus epithelial cell volume and CSF secretion/clearance. BCAS stimulates dynamic changes of TJ proteins and the SPAK-NKCC1 complex, which contributes to altered ChP TJ proteins and infiltration of immune cells. Inflammatory signals activate the IκB kinase (IKK) complex, leading to the phosphorylation and dissociation of IκB proteins (IκBα/β/ε) that normally sequester NF-κB dimers (p65/p50) in the cytoplasm. This process allows NF-κB to move from the cytosol to the nucleus, where it promotes the transcription of target genes like SPAK and NKCC1. Pharmacological inhibition of SPAK with its potent inhibitor ZT-1a provides protective effects via attenuating BCAS-induced structural changes at the ChP and neuroinflammation
留言 (0)