
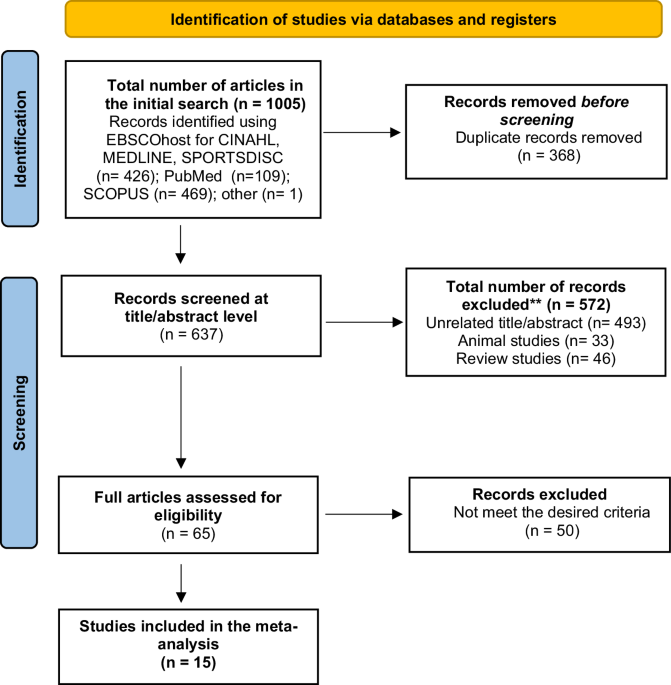
記住我


Two-sample univariate MR analyses were performed to assess the effect between genetically predicted fasting insulin and IGF-1 on childhood and adult obesity. We conducted the analyses on 444,345 European individuals from the UK Biobank participants with self-reported relative childhood adiposity [13]. We verified any findings in a smaller sample of 39,725 participants with objectively measured childhood BMI from the Early Growth Genetics (EGG) consortium [14]. Additionally, we tested the associations with adult BMI using data from 456,426 European individuals from the UK Biobank [15].
MR utilizes GWAS summary statistics data to test the likely causal association between an exposure and outcome of interest. It relies on three key assumptions: (a) the instrumental variants are associated with the exposure; (b) the instrumental variants are not associated with confounders; and (c) the instrumental variants only influence the outcome via the exposure of interest. We selected as instrumental variants those single nucleotide polymorphisms (SNPs) that reached genome-wide significant threshold (P < 5.0 × 10−8) for circulating fasting insulin or IGF-1 concentrations. In all analyses, the variants were aligned to designate the exposure-increasing allele as the effect allele. For variants that were not present in the outcome data, a highly correlated proxy was selected (within 1 Mb and r2 > 0.7; Supplementary Table 1) using a linkage-disequilibrium panel from a random subsample of individuals in the UK Biobank study with self-identified and genetically confirmed white European ancestry.
MR assumes a consistent and linear relationship, but a higher biomarker level may indicate enhanced biomarker action or increased biomarker resistance. To distinguish between these contrasting effects, we applied ‘biological effect’ filtering in our MR models. This method involved filtering genetic variants based on their individual effect on a third phenotype, which serves as an established indicator of that biomarker’s action.
Instrumental variablesFasting insulinGenetic instruments for fasting insulin concentration (logn pmol/L, in models adjusted for BMI) were based on a European GWAS meta-analysis of approximately 150,000 individuals without diabetes, which reported 43 independent signals [16]. As higher insulin concentrations typically reflect greater insulin resistance [17], they can also reflect greater insulin secretion and bioaction [18]. Therefore, we stratified the insulin genetic instruments based on their effect on fasting glucose concentration, as the established role of insulin is to lower circulating glucose [16] (Fig. 1). Genetic data for fasting glucose (BMI adjusted) was based on a European GWAS meta-analysis of 200,621 individuals [16].
Fig. 1: Stratification of biomarker-associated genetic instruments by their biological effects on established downstream traits.
A Fasting insulin associated genetic instruments were filtered by their direction of effect on fasting glucose. B IGF-1 associated genetic instruments were filtered by their direction of effect on childhood height. IGF-1 insulin-like growth factor-1, SNPs single nucleotide polymorphisms.
Of the 43 insulin-increasing alleles, 35 were also directionally associated with higher fasting glucose (indicating insulin resistance), while 7 were directionally related to lower fasting glucose (indicating greater insulin secretion and bioaction). For the remaining one fasting insulin associated variants, there was no available proxy.
IGF-1Genetic instruments for circulating IGF-1 concentration (nmol/L) were based on a European GWAS of 428,525 European participants from the UK Biobank, which we resolved to 828 independent signals [19]. Higher IGF-1 concentrations typically reflect higher IGF-1 secretion and bioaction; but can also reflect IGF-1 resistance (for example, due to damaging mutations in the IGF-1 receptor gene, IGF1R [20]). Therefore, we stratified the IGF-1 genetic instruments by their effect on childhood height, as the established role of IGF-1 is to increase height [11] (Fig. 1). GWAS data for childhood height at age 7 was based on 36,102 children from the Norwegian Mother, Father and Child Cohort Study (MoBa) [21].
Of the 828 IGF-1-increasing alleles, 351 were also directionally related to taller childhood height (indicating greater IGF-1 secretion and bioaction), while 306 were directionally related to shorter childhood height (indicating IGF-1 resistance). For the remaining 171 IGF-1 associated variants, there were no suitable proxies (r2 > 0.7) for childhood height in the MoBa data.
Outcome variablesRelative childhood adiposityGWAS data for relative childhood adiposity in the UK Biobank study were modelled as a primary outcome. This was assessed in 444,345 European participants in response to the question “When you were 10 years old, compared to average would you describe yourself as thinner, plumper, or about average?” (UK Biobank field 1687). The GWAS model treated responses as a linear variable (thinner/average/plumper) [13].
To provide confirmation of the results obtained using relative childhood adiposity, we also used data on measured childhood BMI (in standardized units) from a much smaller GWAS meta-analysis. These data from the EGG consortium were based on a trans-ancestral meta-analysis GWAS of BMI in 39,620 children aged between 2 and 18 years [14].
Adult BMIGWAS data for adult BMI was based on 456,426 European-ancestry participants from the UK Biobank using data from the first assessment centre visit (UK Biobank field 21001) [15].
Statistical analysisMR modelsOur primary analysis was the inverse-variance weighted (IVW) MR model with biological effect and Steiger filtering. Steiger filtering reduces reverse causality by removing instrumental variable SNPs if they have a stronger effect on the outcome than on the exposure (Supplementary Table 2). We performed sensitivity analyses that control for genetic pleiotropy, including MR-Egger, Weighted Median (WM), and Penalized Weighted Median (PWM) MR models. Additionally, we used the MR-Egger intercept P < 0.05, I2 statistic, Cochran’s Q-derived P value, funnel and dosage plots to assess evidence of balanced and unbalanced pleiotropy. When MR-Egger intercept is significant (P < 0.05), we considered the IVW model to be invalid. All analyses were performed using R (version 3.5.1) and a P < 0.0125 was considered statistically significant (calculated as 0.05 / (2 exposures * 2 outcomes)).
留言 (0)