
記住我


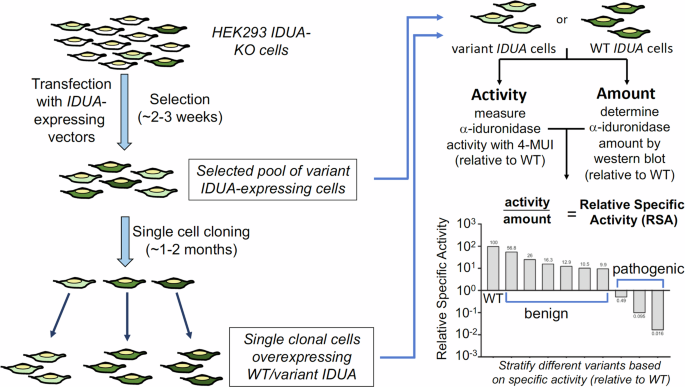
A schematic illustration of the identification of megakaryocyte- and erythroblast-specific differentially methylated regions (DMRs) and the design of ddPCR assays is shown in Fig. 1. First, the genome-wide methylation profiles of erythroblasts, megakaryocytes and other types of blood cells (neutrophils, B cells, and T cells) and tissues (adipose tissues, adrenal gland, colon, esophagus, liver, lung, pancreas and small intestines) were first retrieved from public datasets available on the Roadmap Epigenomics16, the Encyclopedia of DNA Elements (ENCODE)17 project and Blueprint Epigenome18. The methylation densities, defined as the CpG loci being methylated, of all CpG sites on autosomes were systematically compared to identify the DMRs. Details of the retrieval of reference methylomes and data processing for the development of DMRs were described in the Methods section. The megakaryocyte- and erythroblast-specific DMRs were determined using the following criteria:
1.In defining the megakaryocyte-specific and erythroblast-specific DMRs, we would identify a hypomethylated region specific to the target cell types (i.e. megakaryocytes or erythroblasts), in which the methylation densities of the CpG sites in all the other cell or tissue types over the CpG sites in that region exceeded 75%.
2.Multiple CpG sites would be included in the DMR if possible, to improve the signal-to-noise ratio. Also, the selected CpG sites that were located within close genomic proximity would have the comparative advantage for the analysis in plasma DNA molecules, given that plasma DNA is short in nature with a modal size of about 160 bp2. In addition, to facilitate the probe design, DMR with at least 3 CpG sites within 20 bp would be considered.
Fig. 1: Schematic illustration of the detection of megakaryocytic and erythroid DNA through identification of cell-specific markers and digital PCR.
The reference methylomes of various tissues and cells types were studied to identify the megakaryocyte- and erythroblast-specific differentially methylated regions (DMRs). We then developed a digital PCR assay to quantify the methylated and unmethylated DNA sequences each over the megakaryocyte- and erythroblast-specific DMRs, in order to infer the proportions of megakaryocytic DNA and erythroid DNA.
Megakaryocyte- and erythroblast-specific methylation markersBased on the above-mentioned criteria, we have identified a number of DMRs across the whole genome and selected one megakaryocyte-specific DMR and one erythroblast-specific DMR with the largest difference in the methylation densities between the target cell type versus the other cell types (Supplementary Fig. 1). Other candidate DMRs are shown in Supplementary Fig. 2. There is a substantial difference in the methylation levels between erythroblast and megakaryocyte over these two selected DMRs to avoid the cross detection of erythroid and megakaryocytic DNA. The megakaryocyte-specific methylation marker region, named as Mk-1, is located on the paralemmin 2 and A-kinase anchoring protein 2 (PALM2AKAP2) gene on chromosome 9 (Supplementary Fig. 1a). A-kinase anchoring proteins have been found in platelets, but their roles in platelet activation are largely unknown19. The erythroblast-specific methylation marker has been reported by us before and named as Ery-19. The Ery-1 marker is located on chromosome 12 and the genomic region associated with this DMR has not been identified with any annotated gene9 (Supplementary Fig. 1b).
In addition, we have reviewed the methylation densities of megakaryocytes, erythroblasts and other cells and tissues types in the previously reported erythroid methylation marker located in the intronic region of the ferrochelatase (FECH) gene9. As shown in Supplementary Fig. 3a, the 4 CpG sites within this marker were hypomethylated (less than 20%) for both megakaryocytes and erythroblasts. The other previously reported erythroid marker Ery-29, similar to Ery-1, was shown to be specific to erythroblasts when only erythroblasts but not megakaryocytes or other tissue or cell types (except adrenal gland) were hypomethylated in this DMR (Supplementary Fig. 3b).
Detection of megakaryocytic and erythroid DNA through methylation analysis by digital PCRTo deduce the proportions of megakaryocytic and erythroid DNA in plasma through methylation, we have developed a digital PCR assay to quantify the methylated and unmethylated DNA sequences each over the megakaryocyte- and erythroblast-specific DMRs (Fig. 1). Details of the primers and probes of both digital PCR assays are described in the Methods section and Supplementary Table 1. The probes were designed to target fully methylated (all 3 CpG sites being methylated) and fully unmethylated (all 3 CpG sites being unmethylated) DNA sequences over these DMRs after bisulfite treatment of plasma samples. The analytical specificity of these assays was confirmed with the use of synthetic fully methylated and fully unmethylated DNA sequences as positive and negative controls and the details were described in both the Methods section and Supplementary Fig. 4.
Since we applied digital PCR for analysis of methylation in the corresponding DMRs to infer the proportions of megakaryocytic DNA and erythroid DNA, we first studied the relative fractions of unmethylated and methylated DNA haplotypes in the reference methylome data of the different cell types. First, we performed an experiment to determine the reactivity between DNA sequences bearing all the 8 different methylation patterns of the 3 target CpG sites (i.e. -U-U-U-, -M-M-M- and 6 mosaic patterns including -U-U-M-, -U-M-U-, -M-U-U-, -M-M-U-, -M-U-M-, -U-M-M-, where U denotes unmethylated CpG and M denotes methylated CpG) and the probes (which were designed to target fully methylated (-M-M-M-) and fully unmethylated (-U-U-U-) sequences). The results showed that, in the Mk-1-based digital PCR analysis, none of the DNA of mosaic methylation patterns yielded a positive signal above the threshold (Supplementary Fig. 5). Therefore, the subsequent calculation of megakaryocytic DNA contribution would be adjusted according to the relative fractions of fully unmethylated (-U-U-U-) haplotypes versus fully methylated (-M-M-M-) haplotypes only in the different cell types (Fig. 2a). In the Ery-1 marker-based digital PCR analysis, DNA sequences of the -U-U-M- and -M-U-U- patterns also yielded positive signals (i.e. above the pre-defined threshold) with the probe targeting fully unmethylated (-U-U-U-) DNA (Supplementary Fig. 6). Therefore, the subsequent calculation of erythroid DNA contribution would be adjusted according to the relative fractions of -U-U-M-, -M-U-U- and -U-U-U- haplotypes versus fully methylated (-M-M-M-) haplotypes in the different cell types (Fig. 2b). The details of the experiment were described in the Methods section.
Fig. 2: Relative fractions of unmethylated haplotypes and methylated haplotypes in the differentially methylated regions (DMRs) of megakaryocytes and erythroblasts for the adjustment of the calculation of megakaryocytic and erythroid DNA contributions.
The relative fractions in the different cell types are shown. The unmethylated CpG is denoted as U and the methylated CpG is denoted as M. a Megakaryocyte-specific DMR. The relative fractions of the -U-U-U- and -M-M-M- haplotypes in the different cell types are shown. b Erythroblast-specific DMR. The relative fractions of the -U-U-U-, -U-U-M-, -M-U-U- and -M-M-M- haplotypes in the different cell types are shown.
For the megakaryocytic-specific DMR, proportion of unmethylated DNA sequences measured by the corresponding assay would include predominantly proportion of unmethylated DNA from megakaryocytes and a minority from other cell types. Similarly, for the erythroblast-specific DMR, proportion of unmethylated DNA sequences measured by this assay would include predominantly that from erythroblasts and also a minority from other cell types or tissues. Therefore, the proportions of megakaryocytic (MK%) and erythroid DNA (Ery%) in a biological sample would then be derived as:
Let MK% be the proportion of megakaryocytic DNA in plasma,
$$\beginProportion\,of\,unmethylated\,DNA\,sequences\,in\,the\,megakaryocytic\,DMR\,\\ measured\,by\,digital\,PCR\,analysis =Proportion\,of\,unmethylated\,megakaryocytic\,DNA\\\,+Proportion\,of\,unmethylated\,DNA\,contributed\,by\,other\,cell\,types\\ =MK \% \,\times \,relative\,fraction\,of\,fully\,unmethylated\,haplotype\,\\ over\,fully\,unmethylated\,and\,methylated\,haplotypes\,for\,megakaryocytes\,in\,the\,DMR\\ +(1-MK \% )\,\times mean\,relative\,fraction\,of\,fully\,unmethylated\\ \,haplotype\,over\,fully\,unmethylated\\ \,and\,methylated\,haplotypes\,for\,other\,cell\,types\,in\,the\,DMR,\end$$
where the fraction of fully unmethylated DNA haplotype (i.e. -U-U-U-) over fully unmethylated (i.e. -U-U-U-) and methylated (i.e. -M-M-M-) haplotypes of megakaryocyte and the fractions of other cell types (the mean fraction for the other 12 cell types were used in the calculation) within this megakaryocytic DMR (Fig. 2a) were determined based on the reference methylome data and described in the Methods section.
Similarly, let Ery% be the proportion of erythroid DNA,
$$\beginProportion\,of\,unmethylated\,DNA\,sequences\,in\,the\,erythroblastic\,DMR\,\\ measured\,by\,digital\,PCR\,analysis\\ =Proportion\,of\,unmethylated\,erythroid\,DNA\\+Proportion\,of\,unmethylated\,DNA\,contributed\,by\,other\,cell\,types\\ =Ery \% \,\times \,relative\,fraction\,of\,unmethylated\,}\,\\ over\,unmethylated\,and\,fully\,methylated\,haplotypes\\ for\,erythroblasts\,in\,the\,DMR\\ +(1-Ery \% )\,\times \,mean\,relative\,fraction\,of\,unmethylated\,haplotypes\,\\ over\,unmethylated\,and\,fully\,methylated\,haplotypes\,for\,other\,cell\,types\,in\,the\,DMR,\end$$
where the relative fraction of unmethylated DNA haplotypes (including -U-U-U-, -U-U-M- and -M-U-U-) over unmethylated and fully methylated (i.e. -M-M-M-) haplotypes of erythroblast and the fractions of other cell types (the mean fraction for the other 12 cell types were used in the calculation) within this erythroblastic DMR (Fig. 2b) were determined based on the reference methylome data and described in the Methods section.
Detection of plasma erythroid and megakaryocytic DNA in healthy subjectsWe simultaneously performed digital PCR analyses targeting the Mk-1 methylation marker and the Ery-1 marker on plasma samples from 55 healthy subjects. The median age was 57 years old (range: 16–70). The detailed demographic data of the healthy subjects were shown in Supplementary Table 2. Through the digital PCR analysis, the median proportion of megakaryocytic DNA was 44.2% (interquartile range: 37.4%–58.9%). The median proportion of erythroid DNA was 6.2% (interquartile range: 4.3%–8.8%) (Fig. 3). The results suggested that megakaryocytic DNA contributed a substantial proportion to the plasma DNA pool in healthy individuals. We did not observe any correlation between the megakaryocytic DNA proportions and age (r = −0.13, P = 0.35, Pearson correlation), or erythroid DNA and age (r = 0.1, P = 0.46, Pearson correlation) (Supplementary Fig. 7). There was no difference in the proportions of megakaryocytic and erythroid DNA between the male and female control subjects (P = 0.28 for megakaryocytic DNA and P = 0.55 for erythroid DNA) (Supplementary Fig. 8).
Fig. 3: Proportions of megakaryocytic and erythroid DNA in the plasma of healthy subjects.
Each dot represents the proportions of megakaryocytic and erythroid DNA in the healthy subjects. The lower and upper box limits indicate the 25th and 75th percentiles.
In addition, we have studied if there exists any correlation between the proportions of megakaryocytic DNA or erythroid DNA measured in plasma and those measured in buffy coat. We measured the proportions of megakaryocytic and erythroid DNA in the paired buffy coat samples of some (18 out of 55) healthy subjects (those with available samples). The proportion of megakaryocytic DNA measured in buffy coat DNA (median: 1.2%; interquartile range: 0.8%–1.7%) was significantly lower than that in plasma (median: 48.2%; interquartile range: 39.3% – 62.1%) (P < 0.0001, Mann-Whitney U test) among the healthy individuals (Fig. 4a). Similarly, the proportion of erythroid DNA measured in buffy coat DNA (median: 1.1%; interquartile range: 0.8%–1.3%) was also significantly lower than that measured in plasma (median: 4.8%; interquartile range: 3.7%–8.0%) (P < 0.0001, Mann-Whitney U test) (Fig. 4b). No correlation was found between the proportion of megakaryocytic DNA in plasma and that in buffy coat (r = −0.13, P = 0.62, Pearson correlation) (Fig. 4c), or between the proportion of erythroid DNA in plasma and that in buffy coat (r = 0.026, P = 0.92, Pearson correlation) (Fig. 4d). These findings suggested that both the circulating megakaryocytic and erythroid DNA molecules in plasma were unlikely to be derived from circulating erythroblasts and megakaryocytes in the peripheral blood.
Fig. 4: Proportions of megakaryocytic and erythroid DNA in the paired plasma and buffy coat of healthy subjects.
a Proportions of megakaryocytic DNA in the paired plasma and buffy coat samples of healthy subjects. b Proportions of erythroid DNA in the paired plasma and buffy coat samples of healthy subjects. c Correlation between the proportions of megakaryocytic DNA detected in the plasma and the proportions in the paired buffy coat samples of healthy subjects. No correlation was observed. d Correlation between the proportions of erythroid DNA detected in the plasma and the proportions in the paired buffy coat samples of healthy subjects. No correlation was observed.
Plasma DNA analysis in β-thalassemia majorWe performed digital PCR that targeted the Ery-1 marker in plasma of 27 patients with β-thalassemia major. The detailed demographic data of these patients were shown in Supplementary Table 2. Patients with β-thalassemia had significantly higher concentration of total plasma DNA (as measured by Qubit Fluorometer, ThermoFisher) than the control subjects (P < 0.0001, Mann-Whitney U test) (Supplementary Fig. 9). Among these patients with β-thalassemia major, the median proportion of erythroid DNA was shown to be 50.9% (interquartile range: 42.1%–73.3%), which was significantly higher than that among the healthy subjects (median: 6.2%; interquartile: 4.3%–8.8%) (P < 0.0001, Mann-Whitney U test) (Fig. 5a). The erythroid DNA detected in plasma is hypothesised to be derived from erythroblasts in the bone marrow. With such a hypothesis, the observation of higher proportion of erythroid DNA in plasma of β-thalassemia major patients could imply higher number of erythroblasts in the bone marrow. Such result of a higher proportion of erythroid DNA in plasma was compatible with the pathophysiological features of β-thalassemia20 because there is an increased but ineffective erythropoiesis as a result of the imbalance in α- and non-α-globin chains and premature destruction of red cell precursors. Concomitantly, we have analyzed the Mk-1 methylation by digital PCR in these patients. There was no significant difference in the proportion of megakaryocytic DNA deduced by Mk-1 methylation between patients with β-thalassemia major (median: 45.2%; interquartile range: 34.8%–49.7%) and healthy subjects (median: 44.2%; interquartile range: 37.4%–58.9%) (P = 0.46, Mann-Whitney U test) (Fig. 5b).
Fig. 5: Proportions of erythroid and megakaryocytic DNA in the plasma of patients with β-thalassemia major and healthy controls.
a Proportions of erythroid DNA in the patients with β-thalassemia major and healthy subjects. The lower and upper box limits indicate the 25th and 75th percentiles. b Proportions of megakaryocytic DNA in the patients with β-thalassemia major and healthy subjects. The lower and upper box limits indicate the 25th and 75th percentiles.
Plasma DNA analysis in idiopathic thrombocytopenic purpuraWe analyzed the proportion of megakaryocytic DNA in plasma of 10 patients with idiopathic thrombocytopenic purpura (ITP) with the digital PCR assay that targeted the MK-1 methylation marker. The demographic data of the patients were stated in the Supplementary Table 2. All patients had previous bone marrow biopsies showing hyperplasia of megakaryocytes. There was no significant difference in the total plasma DNA concentration (as measured by Qubit Fluorometer, ThermoFisher) between ITP patients and control subjects (P = 0.34, Mann-Whitney U test) (Supplementary Fig. 9). Their median proportion of megakaryocytic DNA (median: 59.9%, interquartile range: 56.7%–64.0%) was significantly higher than that detected among the healthy subjects (median: 44.2%; interquartile range: 37.4%–58.9%) (P = 0.03, Mann-Whitney U test) (Fig. 6a). The higher proportion of megakaryocytic DNA in plasma of patients with ITP is consistent with the compensatory mechanism of increased platelet production from megakaryocytes in response to the peripheral destruction of platelets. We have also studied the Ery-1 methylation in the plasma DNA of these patients with ITP. There was no significant difference in the proportion of erythroid DNA between patients with ITP (median:6.3%; interquartile range: 3.6%–7.7%) and healthy subjects (median: 6.2%; interquartile range: 4.3%–8.8%) (P = 0.41, Mann-Whitney U test) (Fig. 6b).
Fig. 6: Proportions of megakaryocytic and erythroid DNA in the plasma of patients with idiopathic thrombocytopenic purpura (ITP) and healthy controls.
a Proportions of megakaryocytic DNA in the patients with ITP and healthy subjects. The lower and upper box limits indicate the 25th and 75th percentiles. b Proportions of erythroid DNA in the patients with ITP and healthy subjects. The lower and upper box limits indicate the 25th and 75th percentiles.
留言 (0)