Trypanosomatids of the class Kinetoplastea are single-celled eukaryotic parasites [1]. While some trypanosomatid species, such as Leishmania, can be transmitted to humans by insect vectors causing considerable morbidity and mortality [2], most trypanosomatids are monoxenous and exclusively parasitize insects [3]. For these parasites, fecal-oral transmission is the most common mode of pathogen spread [4]. This cycle requires an infected host to defecate parasites onto a food source, where they must remain viable long enough to be ingested by their next susceptible host. Once in the host, parasites often accumulate in the hindgut and rectum, adhering to the lining of these tissues by their single flagellum and dividing by binary fission as attached cells [4]. The structure of this flagellar attachment is similar in all trypanosomatids 5, 6.
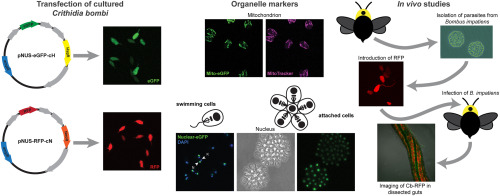
Some trypanosomatids infect bees such as honey bees (Apis mellifera) and bumble bees (Bombus spp.), potentially contributing to pollinator decline [7]. For example, Crithidia bombi is a gut parasite primarily known to infect multiple species of bumble bees, including Bombus impatiens and Bombus terrestris, although C. bombi has recently been found to replicate in the solitary bee species Osmia lignaria and Megachile rotundata as well [8]. In bumble bees, C. bombi impairs learning [9], can reduce queen colony-founding success [10], and can reduce worker survival under stressful conditions [11]. Other trypanosomatids, such as Lotmaria passim, cause similar effects in honey bees [12]. The presence of the parasites triggers an immune response in the bee host, including production of antimicrobial peptides (AMPs), although precisely how infection impacts host fitness is unclear 13, 14. Researchers have also shown that gene expression patterns in cultured parasites differ from those of parasites in the bee gut, representing possible metabolic adaptations to the host environment [14].
Certain floral diets can reduce C. bombi infections in some bumble bee species. The secondary metabolite callunene, discovered in the nectar of heather flowers (Calluna vulgaris), removed or shortened the flagellum of C. bombi and dramatically reduced infection in B. terrestris, presumably by interfering with the parasites’ ability to adhere to and colonize the gut [15]. Similarly, pollen of sunflower (Helianthus annuus) and some other Asteraceae plants dramatically decreases C. bombi infection in B. impatiens 16, 17, 18, 19, 20 but is less effective in other Bombus species [21], suggesting that species-level variation shapes diet-mediated effects on infection outcomes.
The underlying mechanisms for the antiparasitic effect of different floral products such as pollen and nectar are largely unknown. Molecular genetic tools to manipulate parasites for in vivo infections and in culture would facilitate new experimental approaches to understand how floral resources impact host-pathogen dynamics. For instance, which parasite biological processes are disrupted by heather nectar or sunflower pollen? Possible targets include flagellar growth, attachment, and survival and division of attached cells. Discovering the effects of floral products on these activities could improve our understanding of how these different aspects of parasite biology contribute to productive infections. Although all trypanosomatids, including human pathogens, attach to tissues in their insect hosts [6], insect parasites do so in great numbers [4], meaning they could serve as a model for insect colonization by trypanosomatids more generally. In addition, improved understanding of the effects of pollen and nectar diets on the mechanisms underlying parasite infections could allow us to predict the impacts of floral resources on pathogen load and pollinator health.
Detailed study of attachment and modes of cell division would be greatly facilitated by improved visualization of parasites in vivo. For this, both whole cell and organelle markers would allow researchers to monitor the number, location, and cellular structure of parasites at different stages of the infection. Such analyses would improve our understanding of the life cycle of these parasites in their insect hosts, which could reveal vulnerabilities for intervention. In vitro assays for infection behaviors such as attachment would allow for time-resolved, quantitative studies showing how attachment changes under different conditions, predicting infection dynamics in the presence and absence of different floral products and compounds. Finally, genome-wide transcriptomics and proteomics approaches will reveal gene products that mediate interactions between parasites and their insect hosts [13]. Genetic techniques enabling functional knockout and subcellular localization during the cell and life cycle of the parasite would provide important insights into the mechanism of action of specific proteins.
To develop these approaches, our objectives for this study were to 1) establish C. bombi sensitivity to antibiotics used as selectable markers, 2) introduce episomal plasmids including genes for enhanced green fluorescent protein (eGFP) and red fluorescent protein (RFP) into C. bombi cells, 3) create markers for subcellular organelles, 4) isolate and culture parasites from B. impatiens intestinal tracts, and 5) visualize fluorescently-labelled C. bombi cells in vitro and in vivo.

留言 (0)