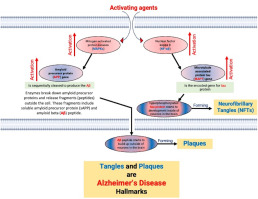
Alzheimer's disease (AD) is an age-related neurodegenerative disorder, accounting for nearly two-thirds of all cases of dementia and up to 20% of individuals older than 80 years of age (Mazure and Swendsen, 2016). With an aging population, AD has been ranked as one of the top ten causes of death (Alzheimers Dement., 2023). >100 years after Alois Alzheimer reporting the first case, amyloid plaques and neurofibrillary tangles (NFTs) remain the main explanation of AD pathogenesis (Zhang, 2023; Zhang et al., 2021). Accordingly, the majority of currently developed therapeutic agents are focusing either one of these two pathological hallmarks (Cummings et al., 2022). An improved strategy could be identifying a drug that is multifunctional or a target that acts on both amyloidogenesis and tauopathy.
Amyloid plaques are composed primarily of abnormally folded Aβ proteins containing either 40 or 42 amino acids. Aβ42 is more abundant than Aβ40 within plaques, attributed to its higher rate of fibrillization and insolubility (Lane et al., 2018). Aβ is generated via a sequential cleavage of the transmembrane protein precursor for amyloid-β (APP) by β-secretase (or BACE1) and γ-secretase, while APP cleavage by the α-secretase A disintegrin and metalloproteinase domain-containing protein 10 (ADAM10) inhibits Aβ production (Sisodia and St George-Hyslop, 2002). In the context of molecular alterations of AD, BACE1 protein is elevated in the brain and body fluids, and connects multiple upstream signaling pathways with downstream amyloidogenesis (Chami and Checler, 2012; Hampel et al., 2021). Conversely, a clue for potential mechanism in amyloidogenesis could be found through a measure of BACE1.
Neurofibrillary tangles are primarily composed of paired helical filaments consisting of hyperphosphorylated tau (Drummond et al., 2020). Abnormal Tau phosphorylation reduces its binding to microtubules and increases its tendency to aggregate, resulting in a loss of its inherent functions. This leads to disruptions in microtubule assembly, axonal transport, dendritic structure, synaptic loss and neuronal death (Hoover et al., 2010; Iqbal et al., 2005; Savelieff et al., 2019). Numerous kinases, including over 20 serine/threonine kinases, have been suggested to phosphorylate Tau (Cavallini et al., 2013; Hanger et al., 2009). Among these kinases, glycogen synthase kinase-3, cyclin-dependent kinase-5 and the MAPK family show strong association between Tau phosphorylation and AD (Iqbal et al., 2005; Johnson and Stoothoff, 2004; Wang et al., 2007).
Staufen-1 (STAU1) is an RNA-binding protein (RBP) containing four double-stranded RNA-binding domains (dsRBDs) (Marión et al., 1999; Wickham et al., 1999). STAU1 gene encodes two major isoforms (63 kDa and 55 kDa) generated by alternative pre-mRNA splicing and/or polyadenylation, which are equally functional (Duchaîne et al., 2000; Park and Maquat, 2013). STAU1 plays an important role in RNA processing including staufen-mediated mRNA decay (SMD) and stress granule formation (Kim et al., 2005; Thomas et al., 2005). These mechanisms mediate the function of STAU1 in cancer development, viral replication and pro-inflammatory effects (Boulay et al., 2014; Gonzalez Quesada and DesGroseillers, 2022; Marcellus et al., 2021; Ye et al., 2019). In the nervous system, STAU1 is shown to regulate neuronal differentiation (Oh et al., 2018), and is involved in the pathogenesis of neurodegenerative disorders including spinocerebellar ataxia, amyotrophic lateral sclerosis and frontotemporal lobar degeneration (Paul et al., 2021; Yu et al., 2012). Interestingly, STAU1 is overexpressed in disease models of AD (Gandelman et al., 2020; Paul et al., 2021), raising a possibility that STAU1 might be associated with amyloidogenesis or tauopathy.
In this study, we provide evidence that STAU1 expression is augmented in cellular and animal models of AD, and define that STAU1 controls amyloidogenesis via the 3’ UTR of BACE1. Through further transcriptome analysis, we find that STAU1 activates P38 MAPK signaling through growth arrest and DNA damage 45 β (GADD45B), which contributes to STAU1-mediated enhancement of Tau phosphorylation at selective sites. Our study highlights a novel role of STAU1 in regulating both amyloidogenesis and tauopathy.



留言 (0)