
記住我

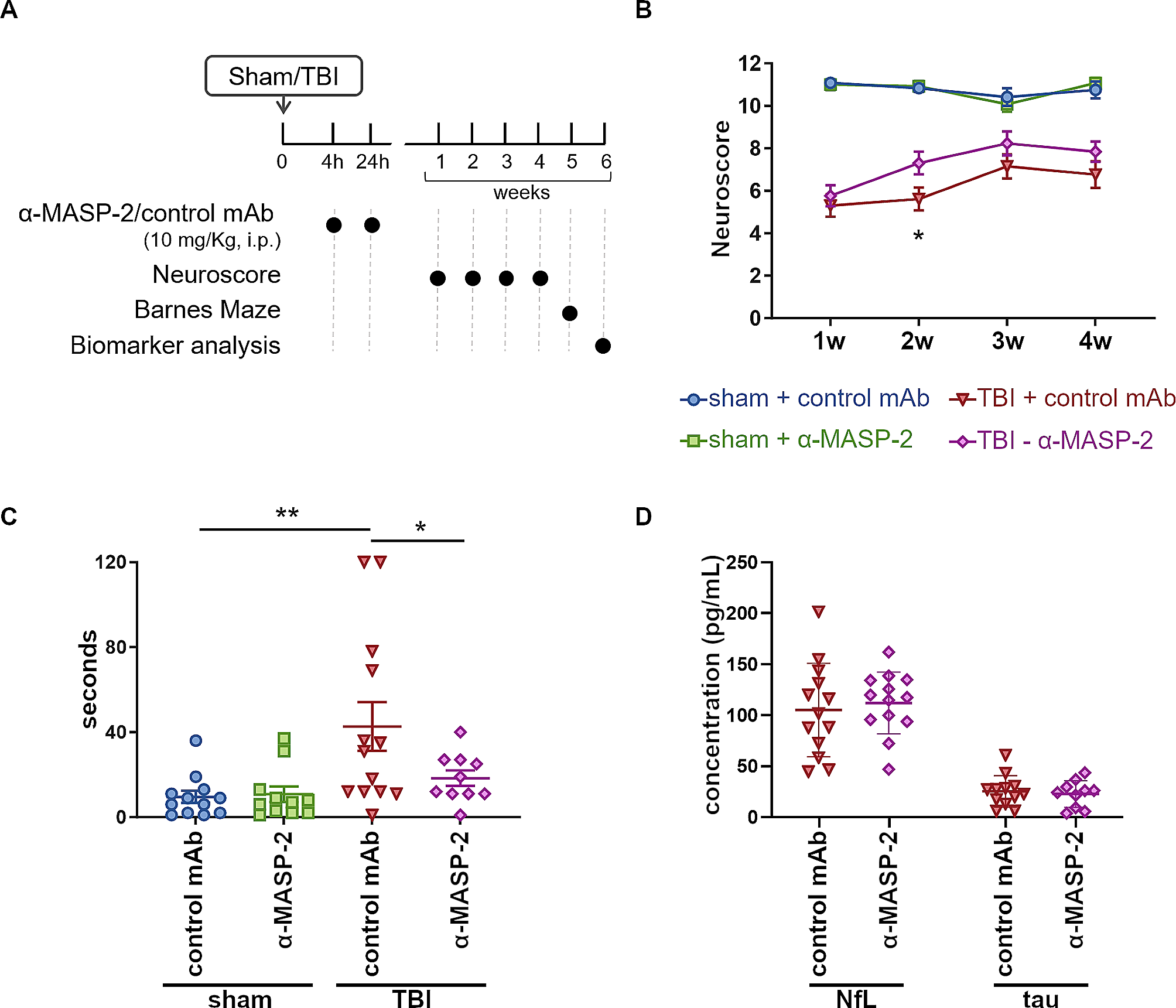





We first conducted a series of pilot studies to determine the optimal dose of K. pneumoniae via intratracheal administration in naïve male and female C56Bl/6J mice (Fig. 1a) and evaluated changes in body weight (Fig. 1b), lung bacterial load (Fig. 1c) and mortality (Fig. 1d), alongside observable symptoms of illness. A low dose of 1 × 104 CFU failed to induce any observable symptoms in inoculated mice, and a negligible bacterial load in the lungs at 24 h (< 3 CFU/lung; n = 3; not shown). Mice that received a dose of 1 × 105 CFU had a low bacterial load, and exhibited only minimal symptoms (inactive). The intermediate dose of 1 × 106 CFU induced overt clinical symptoms of illness acutely (e.g., inactive, fur erect, hunched posture, acute dyspnea), and an intermediate lung bacterial load. Mice that received the highest dose of 1 × 107 CFU were found to have a high lung bacterial load, exhibited pronounced weight loss within 48 h, and presented with significant clinical symptoms of acute respiratory illness. This dose also resulted in considerable mortality (63%; n = 5/8) within the first week post-infection (2/4 males and 3/4 females), between 2 and 3 days post-inoculation, as animals reached the weight loss threshold requiring humane euthanasia. No mortality was observed for mice that received a dose of 1 × 106 CFU K. pneumoniae or vehicle (Fig. 1b).
In a separate pilot experiment, we compared intratracheal treatments of sterile saline only versus broth (the vehicle for K. pneumoniae inoculation), and found no mortality, comparable body weights, no signs of acute illness, and no differences in lung histological measures (n = 4/group; not shown). Subsequent experiments therefore employed a broth solution vehicle or a dose of 1 × 106 CFU K. pneumoniae.
K. pneumoniae administration after TBI does not worsen acute symptoms of illnessTo assess whether K. pneumoniae infection after TBI altered functional outcomes, mice were subjected to TBI or sham surgery, followed by K. pneumoniae or vehicle inoculation at 4 days post-injury, and examined over the subsequent 7 d period (Fig. 2a). The lung bacterial load at 24 h post-infection was comparable between Sham-Kp and TBI-Kp mice (t8 = 0.62, p = 0.5529; Fig. 2b), and undetectable in vehicle-treated mice. Spleens were collected fresh and weighed at either 24 h or 7 d post-infection, as a surrogate indication of mobilization of the peripheral immune response. However, neither TBI nor K. pneumoniae altered spleen weight (either raw or relative to body weight) (not shown; p > 0.05).
Fig. 2
K. pneumoniae infection causes transient hypoactivity and weight loss in both TBI and Sham mice. The experimental timeline (a), whereby TBI or sham mice were inoculated with Kp or vehicle after 4 days, then were assessed repeatedly at 4 h, 24 h and 7 d post-infection (OF = Open Field behavior test). At 24 h post-infection, the lung bacterial load was equivalent in sham and TBI mice (b). Changes in body weight across the 7 d time course (c; n = 10/group) revealed a main effect of time, and a time-x-Kp interaction (*p < 0.05 from post-hoc analyses, 3-way RM ANOVA). In the OF test at 4 h post-infection, a non-significant trend towards reduced time in the center was observed in Kp-treated mice (d), which was resolved by 24 h and 7 d (not shown). Also in the OF test, TBI induced an increase in activity at 4 h, 24 h and 7 d (e-g; 2-way ANOVAs, main effect of TBI *p < 0.05, ***p < 0.001), while Kp independently reduced activity just at 4 h post-infection (e; ****p < 0.0001). n’s in (d–g) are depicted by individual data points; males indicated by closed circles/diamonds and females indicated by open circles/diamonds
Body weights were assessed daily as an indicator of general health. A main effect of time was observed (F11, 396=21.30, p < 0.0001), as well as a time-x-Kp interaction (F11, 396=3.75, p < 0.0001). All groups appeared to exhibit comparable mild weight loss (approximately 2–4%) in the first 24 h post-TBI/sham surgery then quickly regained it, although this change was not statistically significant. Then, at 24 h (1 day) post-inoculation, both sham-Kp and TBI-Kp groups showed a similar reduction in body weight compared to vehicle-treated groups (post-hoc *p < 0.01 compared to Sham-Vehicle); although again this was only a transient drop. When cohorts were separated by sex (n = 3/group; Supplementary Fig. 1), it was noted that males showed more consistent body weights across the time course; whereas females showed considerable within-group variability. The effect of Kp on body weight was more pronounced in female mice, as a time-x-Kp interaction was detected in females (F11, 176=2.39, p = 0.0087) but not males (F11, 176=1.53, p = 0.1244).
The open field test was conducted to evaluate locomotor activity and anxiety-like behavior. At 4 h post-infection, a non-significant trend was seen with K. pneumoniae-treated mice appearing to spend less time in the center of the arena (F1, 35=3.72, p = 0.0619), suggestive of increased anxiety-like behavior (Fig. 2d). At the same time, independent main effects of both TBI (F1, 35=61.62, p = 0.0255) and K. pneumoniae (F1, 35=5.45, p < 0.0001) were observed for distance traveled in the open field, with TBI mice moving more but K. pneumoniae-treated mice moving less (Fig. 2e). This hypoactive effect of K. pneumoniae was transient, as it had resolved by the 24 h (Fig. 2f) and 7 d (Fig. 2g) time points. However, a main effect of TBI was observed across the time course (F1, 59=6.38, p = 0.0143 at 24 h, and F1, 36=13.67, p = 0.0007 at 7 d).
Elevated serum inflammatory cytokines in response to K. pneumoniaeFor prototypical pro-inflammatory cytokine IL-1β (Fig. 3a), a significant TBI-x-Kp interaction was detected (F1, 16=5.95, p = 0.0267), whereby IL-1β was elevated in the serum of TBI-Kp mice compared to TBI-vehicle (Tukey’s post-hoc, p = 0.0397). Independent of TBI, K. pneumoniae induced a robust increase in cytokines IL-6 (Fig. 3b; F1, 15=10.01, p = 0.0064) and TNFα (Fig. 3c; F1, 15=15.86, p = 0.0011), as well as the chemokines CCL2 (Fig. 3e; F1, 16=7.15, p = 0.0167), CXCL1 (Fig. 3f; F1, 15=7.21, p = 0.0163) and CXCL2 (Fig. 3g; F1, 16=17.29, p = 0.0010). By contrast, levels of anti-inflammatory cytokine IL-10 were not altered by K. pneumoniae or TBI (Fig. 3d).
Fig. 3
Elevated Serum Cytokines at 24 h Post-Infection. Cytokine levels in serum at 24 h post-infection (a–g), reveal a pro-inflammatory effect of K. pneumoniae, but no influence of a prior TBI on this response (2-way ANOVAs, main effects of Kp *p < 0.05, **p < 0.01). In (a), *p < 0.05 indicates Tukey’s post-hoc comparison (TBI-vehicle vs. TBI-Kp) following a significant TBI-x-Kp interaction. Males denoted by closed circles/diamonds and females by open circles/diamonds. n = 4–5/group
Lung inflammation and pathology after K. pneumoniae infectionPulmonary inflammation and lung pathology was evaluated in a subset of animals at 24 h and 7 d post-K. pneumoniae infection, to evaluate whether the combined insult of TBI plus K. pneumoniae worsened pulmonary outcomes. At 24 h post-infection, when the presence of bacteria was confirmed in lung homogenates (Fig. 2b), the analysis of inflammatory cytokines in BALF revealed a robust pro-inflammatory response in K. pneumoniae-treated mice compared to vehicle. Specifically, K. pneumoniae induced a robust increase in cytokines IL-1β (Fig. 4a; F1, 15=4.66, p = 0.0476), IL-6 (Fig. 4b; F1, 15=27.99, p = 0.0001), TNFα (Fig. 4d; F1, 15=27.82, p < 0.0001) and IL-10 (Fig. 4e), as well as the chemokines CCL2 (Fig. 4c; F1, 15=13.43, p = 0.0023), CXCL2 (Fig. 4f; F1, 15=13.66, p = 0.0022) and CXCL1 (Fig. 4h; F1, 16=12.53, p = 0.0027). However, a TBI sustained 4 days prior to K. pneumoniae infection did not alter BALF cytokine levels.
Fig. 4
K. pneumoniae Infection Alters Lung Histology and BALF Cytokine Levels. Cytokine levels in bronchoalveolar fluid (BALF) at 24 h post-infection (a–f, h) reveal a predominantly pro-inflammatory effect of K. pneumoniae but no effect of prior TBI (2-way ANOVAs, main effect of Kp; *p < 0.05; **p < 0.01; ****p < 0.0001). Immunostaining for MPO + neutrophils (colocalized with nuclear marker DAPI; g) in lung tissue revealed a robust neutrophilic infiltration at 24 h post-infection, particularly associated with vascular thrombi (*). By 7 d post-infection, this immune response had resolved, with no differences in MPO + cell numbers between the groups (i). Representative H&E staining illustrates lung histology at 7 d post-infection (j). Quantification of the mean linear intercept (MLI) (k), where an increase reflects pathology, revealed an increase in lungs from TBI-Kp mice (2-way ANOVA, TBI-x-Kp interaction; Tukey’s post-hoc *p < 0.05). Males denoted by closed circles/diamonds and females by open circles/diamonds. n = 4–6/group. Scale bar = 50 μm (g) and 100 μm (j)
Significant correlations between serum and BALF levels of several cytokines were noted, for IL-1β (r2 = 0.50, p = 0.0007), TNFα (r2 = 0.32, p = 0.0121), IL-6 (r2 = 0.43, p = 0.0024), CXCL1 (r2 = 0.63, p < 0.0001), CXCL2 (r2 = 0.25, p = 0.0256) and IL-10 (r2 = 0.38, p = 0.0047), and a trend observed for CCL2 (r2 = 0.20, p = 0.0559), suggesting that the local pulmonary response was the source of inflammatory mediators in the serum. Considerable neutrophil infiltration was observed by MPO immunofluorescence staining of lung tissue at 24 h, particularly associated with vascular thrombosis (Fig. 4g). Qualitatively this infiltration appeared comparable between Sham-Kp and TBI-Kp groups, although quantification was not readily possible due to the dense clustering of these cells at this time.
By 7 d post-infection, fewer MPO + cells were detected in lung tissue (Fig. 4i) and quantification revealed similar numbers across all groups (n.s.). Also at 7 d post-infection, calculation of the mean linear intercept (MLI) to evaluate lung pathology from H&E-stained sections (Fig. 4j) revealed a significant TBI-x-Kp interaction (F1, 20=5.56, p = 0.0287) and a trend towards a main effect of K. pneumoniae (F1, 20=3.71, p = 0.0685). From Tukey’s post-hoc comparisons, K. pneumoniae increased the MLI in TBI mice specifically (TBI-vehicle vs. TBI-Kp; p < 0.05), indicating the presence of pathology (reduced structural integrity of the alveolar septum/increased alveolar size). The TBI-Kp group trended towards a higher MLI compared to Sham-Kp mice (p = 0.0593; Fig. 4k).
Immune-related gene expression changes in the injured brain after k. pneumoniae infectionWe next considered the potential effect of K. pneumoniae lung infection on the brain’s immune response to TBI, by examining the expression profiles of key genes involved in regulating neuroinflammation–including cytokines, chemokines, immune cell markers, and downstream signaling mediators. Broadly, at 24 h post- K. pneumoniae infection (= 5 d post-TBI/sham), most genes exhibited upregulation in response to TBI in the ipsilateral cortex (Fig. 5a) and ipsilateral hippocampus (Fig. 5h). In the cortex, TBI induced increased expression of cytokines Il1β (Fig. 5b; F1, 19=6.91, p = 0.0165) and Tnfα (Fig. 5c; F1, 19=23.03, p = 0.0001), chemokine Ccl2 (Fig. 5d; F1, 19=47.93, p < 0.0001), leukocyte marker Cd45 (Fig. 5e; F1, 19=17.54, p = 0.0005), as well as immune mediators Arg1 (Fig. 5f; F1, 19=27.83, p < 0.0001) and Hmox1 (Fig. 5g; F1, 19=26.20, p < 0.0001). Independently, K. pneumoniae induced an increase in Tnfα only (F1, 19=5.96, p = 0.0246). However, a significant TBI-x-Kp interaction was observed for Ccl2 (F1, 19=7.06, p = 0.0156) and Hmox1 (F1, 19=9.99, p = 0.0051), as well as a non-significant trend towards an interaction for Arg1 expression (F1, 19=3.73, p = 0.0685). From Tukey’s post-hoc analyses, TBI-Kp mice were found to have higher levels of Ccl2 expression compared to either TBI-vehicle or Sham-Kp mice (Fig. 5d; p < 0.05). Arg1 expression was only elevated in TBI-Kp compared to Sham-Kp mice (Fig. 5f); while for Hmox1, expression in TBI-Kp mice was higher compared to both TBI-Vehicle and Sham-Kp mice (Fig. 5g). Altogether, TBI induced a robust pro-inflammatory immune state in the injured cortex, and post-injury K. pneumoniae lung infection appeared to potentiate this response for select genes.
Fig. 5
K. pneumoniae (Kp) infection had a minimal impact on immune-related gene expression in the injured brain. Quantatitive PCR evaluated the relative expression of multiple genes of interest, with group means depicted in heatmaps of the ipsilateral cortex (a) and ipsilateral hippocampus (h) at 24 h post-Kp/vehicle (= 5 d post-TBI/sham). Heatmap key illustrates that increasing color saturation reflects higher gene expression relative to the Sham-vehicle control group (bolded gene names are those that are also depicted graphically; scale = fold change gene expression vs. Sham-Vehicle). Group comparisions for select genes of interest are shown for the cortex (b–g) and hippocampus (i–n), with analysis by 2-way ANOVAs followed by Tukey’s post-hoc comparisons when a significant TBI-x-Kp interaction was observed (i.e., for d, g and f). Males are indicated by closed circles/diamonds and females by open circles/diamonds. n = 4–6/group
In the hippocampus (Fig. 5i-n), examination of the same key genes revealed main effects of TBI (Il1β: F1, 17=4.91, p = 0.0406; Tnfα: F1, 17=5.86, p = 0.0270; Ccl2: F1, 17=15.63, p = 0.0010; Cd45: F1, 17=5.00, p = 0.0391; Arg1: F1, 17=5.76, p = 0.0281; Hmox1: F1, 17=6.21, p = 0.0233). However, no independent effects of K. pneumoniae were observed, nor were there any TBI-x-Kp interactions. The chemokine Cxcl2 was minimally detected in the cortex, but below the threshold for detection in the hippocampus, while the anti-inflammatory cytokine Il-10 was below detection in both brain regions regardless of experimental group.
Cellular neuroinflammation after TBI was not affected by K. pneumoniae infectionAt a sub-acute time point of 7 d post-infection (= 11 d post-injury), markers of inflammatory immune cells were detected by immunofluorescence staining on coronal sections to evaluate whether K. pneumoniae altered their infiltration or activation (Fig. 6a). Quantification of MPO + cells, presumably neutrophils, revealed a TBI-induced increase (Figs. 2 and 6a-way ANOVA F1, 20 = 6.04, p = 0.0232). This response was highly variable, ranging from 0 to 310 in TBI brains. Quantification of GFAP + astrogliosis was performed by determining the proportion of GFAP + coverage of a consistent region-of-interest positioned in the core of the cortical lesion (Fig. 6c) or dorsal hippocampus (Fig. 6d). TBI induced a robust increase in GFAP reactivity at this time in both regions of ∼ 20% compared to ∼ 5% in sham-operated mice (F1, 20 =45.63, p < 0.0001; F1, 20 = 37.22, p < 0.0001), to a similar extent in both vehicle- and K. pneumoniae-treated mice.
Fig. 6
Immunofluorescence staining of cellular neuroinflammation after TBI and K. pneumoniae infection. Representative immunofluorescence images from the ipsilateral (injured) cortex of a TBI-Vehicle mouse at 11 days post-injury (7 d post-vehicle), illustrating detection of GFAP + astrocytes and MPO + neutrophils (a). Scale bar = 100 μm. Quantification of the total number of MPO + cells in the injured hemisphere (b), as well as GFAP % area coverage in the lesioned cortex (c) and hippocampus (d), revealed a main effect of TBI but no effects of K. pneumoniae (2-way ANOVA, *p < 0.05, ****p < 0.0001)
The extent of brain tissue damage after TBI was not affected by K. pneumoniaeAt 7 days post-infection (= 11 days post-TBI), brains were collected, sectioned, and stained with CV and LFB to allow for histopathological assessment (Fig. 7a). Volumetric analysis of the remaining ipsilateral cortex tissue revealed a significant effect of TBI on cortex volume (2-way ANOVA F1, 20=97.85, p < 0.0001; Fig. 7b), as well as loss of hippocampal tissue compared to sham controls (F1, 20=10.55, p = 0.004; Fig. 7c). However, K. pneumoniae and vehicle-treated mice did not differ. Consideration of sex as a variable by 3-way ANOVA revealed a main effect of sex on cortical volume only (F1, 16=6.70, p = 0.0198), but no interactions, indicating that females had a slightly larger ipsilateral : contralateral cortex volume overall.
Fig. 7
K. pneumoniae Infection Did Not Alter Cortical and Hippocampal Neuropathology After TBI. Representative brain sections from each experimental group (a) illustrate the extent of tissue damage from CV/LFB staining. Quantification of remaining intact tissue in the cortex (b) and hippocampus (c), expressed as ipsilateral relative to contralateral regions, demonstrates the considerable loss of tissue as a result of TBI (2-way ANOVA, **p < 0.01; ****p < 0.0001), but no effect of K. pneumoniae nor a TBI-x-Kp interaction. Males are indicated by closed circles and females by open circles. Scale bar = 1000 μm
Fecal bacterial diversity and composition were not altered by TBI or lung infectionThe fecal microbiome was examined, across a time course post-injury and K. pneumoniae infection, as recent evidence supports a complex bi-directional relationship between the brain and the gut in the context of neurotrauma as well as in infection-related immune responses [38,39,40].
Alpha-diversity, as measured using the phylogenetic tree-weighted Hill-Shannon metric, was comparable between injury and treatment groups at all time points (Fig. 8B). Likewise, beta-diversity was similar between groups, and PERMANOVA tests to assess the impact of injury and K. pneumoniae on community structure revealed no significant differences at any time point (Fig. 8C). Fecal samples at all time points showed high abundance of Bacteroides, Roseburia, and Turicibacter (Fig. 8D).
Fig. 8
The Fecal Microbiome was Not Altered by TBI or Lung Infection. (a) Time line of sample collection. (b) Violin plots representing phylogenetic tree-weighted Hill-Shannon alpha-diversity. (c) Principle coordinate analysis (PCoA) plots of the weighted UniFrac distances shown in the first two principal coordinates for bacterial taxonomic composition. Ellipses represent the 90% confidence interval around the group centroid. Results for PERMANOVA testing are shown for injury and treatment differences (SS [sum of squares] shows effect size and R2 shows variance explained). (d) Bacterial genus-level relative abundance data grouped by injury + treatment group and timepoint
At both the ASV and genus level, non-parametric repeated-measures Brunner-Langer tests did not detect any effect of injury, K. pneumoniae, or time on bacterial abundance. Subsequent linear modelling approaches at individual time points to investigate potential group-wise effects also showed no significant differences between groups.
留言 (0)