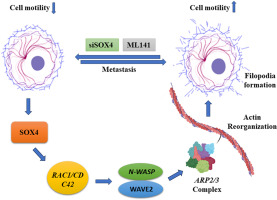
SOX4 induces cytoskeleton remodeling and promotes cell motility via N-WASP/ARP2/3 pathway in colorectal cancer cells
Most cancer-related fatalities in people with CRC are caused by metastasis, which is the process by which cancer cells spread from the primary to a distant site [1]. Tumor cells must effectively complete each stage of “metastatic cascade” in order to generate metastatic lesions: disseminate from the primary tumor → migration & invasion → intravasation → extravasation → subsequent tumor growth at secondary site. Cell movements and changes in shape throughout this intricate process necessitate a substantial spatial and temporal reorganization of the cytoskeleton [2]. In order to induce motility in cancer metastasis, aberrant actin filament assembly produces membrane protrusion. Cancer cells develop a variety of actin-based protrusions during migration and invasion, including lamellipodia, filopodia, invadopodium, peripheral ruffles (PRs), and circular dorsal ruffles (CDRs) [3]. Filopodia are small bundles of actin that contain plasma membrane protrusions and are crucial for numerous physiological functions, including angiogenesis, cell adhesion, cell-cell interactions, and migration. Filopodia are the pioneers at the leading edge of migrating cells that probe the surroundings for signals. Numerous cell types are stimulated to migrate by filopodia, and an increase in filopodia density has been linked to cancer[4]. Actin nucleators like the RAC1/CDC42, ARP2/3 complex, N-WASP family, and formins facilitate membrane protrusions during cell motility by causing the actin filaments to assemble into filopodia and lamellipodia [5]. Through WAVE regulatory complex activation and actin assembly dependent on the ARP2/3 complex, RAC alone can cause the development of protrusions, which is necessary for effective cell migration. Both CDC42 and RAC1 are responsible for the actin modifications in the absence of WAVE regulatory complex. However, these protrusions are less in number, also smaller in size than the canonical[6]. The development of filopodia-like structures in this reconstitution system requires ARP2/3 complex-dependent nucleation.
The SRY-related high mobility group (HMG) box or SOX family of transcription factors play critical roles in normal development including heart, pancreas, hematological, lymphatic, and vascular system [7]. Multiple lines of evidence suggest that the SOX4 gene is an oncogene since it is commonly amplified and overexpressed in more than 20 different forms of cancer. The interactions between SOX4 and other transcription factors make many of its effects on gene expression tissue- and context-specific. Nevertheless, many of the impacts of SOX4 hyperactivity share common characteristics, including the enhancement of cell survival, stemness, the epithelial to mesenchymal transition (EMT), migration, and metastasis [8]. SOX4 along with SOX11 has been shown to regulate genes involved in cytoskeletal/extracellular matrix (ECM) organization. Induction of SOX4 and SOX11 suppresses epidermal terminal differentiation while deficiency enhanced differentiation and impairs cell motility. Fascin actin-bundling protein 1 (FSCN1) was found as a direct target of SOX4, which reactivation led to cell migration [9].
However, little is known about involvement of SOX4 in the precise molecular processes that underlie actin filaments reorganization and cell motility in the development of CRC. Understanding the molecular basis of cell motility is therefore crucial since it could lead to the development of novel therapeutic strategies to prevent metastasis. In this study, we demonstrated that SOX4 involves in filopodia formation, and cell motility through N-WASP/ARP2/3 complex. Overexpression of SOX4 increased cell’s migration and invasion. SOX4 depletion causes the dysregulation of multiple genes involved in cytoskeleton remodeling and metastasis in colorectal cancer (CRC).








留言 (0)