JDM is an acquired idiopathic inflammatory myopathy characterized by symmetric proximal muscle weakness and distinctive cutaneous characteristics [1,2]. Compared with adult dermatomyositis (DM), JDM manifests more frequently as calcinosis cutis, cutaneous vasculitis, ulcerations, and vasculopathy of the gastrointestinal tract [2]. The first-line treatment is glucocorticoids plus methotrexate or cyclosporin [3,4]. For severe and refractory cases, tacrolimus, intravenous immunoglobulin (IVIG), mycophenolate mofetil, rituximab, or antitumour necrosis factor (TNF)-α can be considered [5,6]. We reported the successful use of JAK inhibitors (JAKis) in JDM and evaluated the long-term use of JAKis [7], demonstrating that they could be a suitable and safe choice for JDM treatment with relative safety.
As an important part of the host response, type I IFNs secreted by various immune cells play a powerful antiviral role. However, continuously elevated type I IFN signalling is closely related to the onset and progression of JDM. Previous studies have reported that juvenile and adult DM patient blood cells and muscle tissue displayed higher type I IFN signals, which is considered to be a key feature of DM [8,9]. The powerful effect of JAKis in the treatment of DM is due to their targeting of the IFN-JAK-STAT pathway. Although therapy targeting the IFN pathway has achieved certain effects, relapse and persistent muscle damage are still unresolved, which may be related to specific immune cells with memory functions, such as T and B cells.
The link between adaptive immunity, especially T-cell immunity, and JDM remains largely unclear. IFN can act not only on innate immunity but also on adaptive immunity [10]. IFN-α/β directly influences the fate of CD4+ and CD8+ T cells during the initial phases of antigen recognition [10]. Type I IFNs also drive the expansion of CD8+ T cells during infection with lymphocytic choriomeningitis virus [11]. However, for the memory subgroup, when antigen-specific memory CD8+ T cells are treated with type I IFNs in vitro, they do not exhibit substantial functional activation. This result suggests that type I IFNs require additional secondary signals or accessory cells to fully activate bystander T cells [12]. Evidence has shown that virus-induced type I IFNs rapidly decrease the number of CD8+ T cells, which may accelerate the depletion of effector CD8+ T cells [13]. CD8+ T cells play a key role in autoimmune diseases as a cell with a killing function. In SLE patients with class III or IV nephritis, CD8+ T cells are one of the predominant infiltrating immune cell types, and their accumulation in periglomerular areas correlates with the severity of disease [14]. CD8+ T cells may be critical mediators in the pathogenesis of dermatomyositis. Many studies on experimental myositis (EAM) models have mentioned that deletion of CD8+ T cells prevented the establishment of a typical myositis phenotype induced by antibodies or myosin in mice [15,16]. A recent scRNA-seq analysis confirmed that type I IFN signalling in the periphery could regulate T cells, and the interaction between immune cells and smooth muscle cells (SMCs) or fibroblasts in muscle was enhanced under the inflammatory state [17]. This analysis suggests that T cells affected by type I IFN may be involved in the pathogenesis of JDM in target organs.
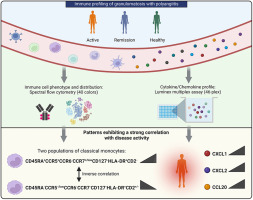
There has been no research on the regulatory effect of elevated type I IFN signalling on CD8+ T cells and the further impact on JDM disease progression. This study aimed to investigate the comprehensive CD8+ T-cell state in active JDM patients, pinpoint the relationship between strong IFN activation and CD8+ T-cell distribution and function, and explore possible targeted therapy against JDM (Fig. 1).










留言 (0)