Acute kidney injury (AKI) is a clinical syndrome characterized by a rapid decline in kidney function [1]. Research indicates that 14–29 million patients suffer from AKI, with a detection rate of 2.03 % by expanded criteria [2]. Over half of intensive care unit (ICU) patients experience AKI, leading to increased serum creatinine levels and the development of end-stage renal diseases [3]. Causes of AKI include sepsis, nephrotoxic drugs, and renal ischemia-reperfusion injury [4]. Sepsis-induced acute kidney injury (S-AKI) accounts for 47.5 % of cases and is the most prevalent type of AKI [5]. Sepsis, a life-threatening organ dysfunction resulting from a dysregulated host response to infection [6], can impact the kidneys through inflammation, hypoxia, and oxidative stress, leading to tubular and renal interstitial damage and ultimately AKI [7]. Patients with S-AKI have a higher incidence and poorer prognosis [8]. However, the precise pathogenesis of S-AKI remains unknown, highlighting the importance of further investigation into the disease mechanism and identification of new therapeutic targets.
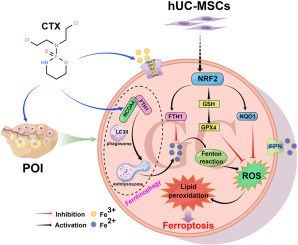
Ferroptosis, characterized by intracellular iron accumulation and lipid peroxidation, represents a newly discovered form of programmed cell death distinct from apoptosis, necrosis, and autophagic cell death in terms of morphology, biochemistry, and genetics [9]. Lipid peroxidation [10] and iron overload [11] are central events in ferroptosis pathogenesis. Various regulatory pathways, including iron and lipid metabolism [9], glutamate-cystine antiporter (System Xc-) [12], and redox imbalance [13], contribute to ferroptosis. Glutathione peroxidase 4 (GPX4) acts as a crucial ferroptosis suppressor protein, maintaining redox homeostasis and protecting cell membranes from lipid peroxide damage by utilizing glutathione (GSH) as a reducing cofactor [14]. Acyl-CoA synthetase long-chain family member 4 (ACSL4) serves as a key execution factor in ferroptosis, promoting the synthesis of phosphatidylethanolamine (PE) species containing arachidonic acid (AA) and adrenic acid (AdA), substrates for lipid peroxidation [15]. Ferroptosis has been observed in folic acid-induced AKI and cisplatin-induced AKI in kidney research [16,17].
Autophagy, a self-repair process involving the removal of excess proteins and organelles through lysosomal degradation [18], encompasses macroautophagy, microautophagy, and chaperone-mediated autophagy (CMA) [19]. Lipid droplets, dynamic organelles found in the endoplasmic reticulum (ER) membrane, consist of neutral lipids like triglycerides and cholesterol [20]. Lipid droplets undergo hydrolysis to release fatty acids (FAs) through lipolysis [21]. Additionally, autophagosome-coated lipid droplets can fuse with lysosomes to release fatty acids, a process known as lipophagy [22]. RAB7, a small guanosine triphosphatase (GTPase), plays a central role in regulating lipophagy in hepatocellular cells by facilitating lysosome recruitment to the lipid droplet (LD) surface [23]. LDs are surrounded by the perilipin (PLIN) family, with perilipin 2 and perilipin 3 being ubiquitously expressed [24]. While lipophagy studies have primarily focused on liver diseases, such as nonalcoholic fatty liver disease (NAFLD) [25] and adipocytes [26], there is limited research on kidney diseases.
Previously, ferroptosis was believed to be distinct from autophagy [9]. However, mounting evidence suggests that autophagy, including lipophagy [27], ferritinophagy [28], and clockophagy [29], plays a crucial role in the onset of ferroptosis. Overactivation of lipophagy can lead to the accumulation of free fatty acids, causing increased cellular lipotoxicity and lipid peroxidation [30]. Inhibiting lipophagy has been shown to alleviate ferroptosis [27,31]. Nonetheless, the precise upstream pathway of lipophagy and its modulation of ferroptosis development in renal tubular epithelial cells, potentially delaying the progression of S-AKI, remain unclear.
Here, this study hypothesizes that lipophagy activation drives ferroptosis progression in renal tubular epithelial cells, with RAB7 serving as a pivotal factor exacerbating S-AKI development. Genetic inhibition of lipophagy through siRNA targeting RAB7 has been found to mitigate ferroptosis and attenuate S-AKI. The objective of this study is to establish a novel theoretical foundation for future S-AKI treatment.
Male C57BL/6 mice, aged 6–8 weeks and weighing 20–25 g, were administered LPS (15 mg/kg, O55:B5, L2880, Sigma‒Aldrich) or PBS through intraperitoneal injection (ip). 30 min early intraperitoneal injection of Fer-1 (5 mg/kg, HY-100579, MedChemExpress), followed by intraperitoneal injection of LPS (15 mg/kg) for the Fer-1 group. The mice were sacrificed after 24 h to harvest serum and kidneys for subsequent analysis. All animal experiments were performed according to the Animal Protocol Committee of Shanghai Jiao Tong University and were approved by the Animal Care Committee of Renji Hospital, School of Medicine, Shanghai Jiao Tong University.
The human renal proximal tubular cell line (HK-2 cell line) was obtained from the American Type Culture Collection (ATCC®, CRL-2190) and cultured in Dulbecco's modified Eagle's medium (DMEM)/F-12 (Thermo Fisher Scientific, 11,330,057) supplemented with 1 % penicillin‒streptomycin (Thermo Fisher Scientific, 10,378,016) and 10 % fetal bovine serum (Thermo Fisher Scientific, 10,099,158). Thirty minutes before LPS administration, HK-2 cells were pretreated with Fer-1 (5 μM, HY-100579, MedChemExpress) followed by LPS (100 μg/ml, O55:B5, L2880, Sigma‒Aldrich) for 24 h as previous studies [32].
Total RNA was extracted from the renal cortex of control and LPS mice by using an RNA kit (Cat. No. ID: 931636, QIAGEN) according to the manufacturer's instructions. RNA sequencing was performed on the Illumina platform (HiSeqTM 2500, Illumina, Shanghai OE Biotech. Co., Ltd.). The genes with P < 0.05 and fold changes <0.5 or >2 were regarded as DEGs.
Serum from mice was obtained when sacrificed after 24 h for the measurement of serum creatinine and BUN for kidney function. The serum creatinine (Nanjing Jiancheng Bioengineering Institute, C011-2-1) and BUN (Nanjing Jiancheng Bioengineering Institute, C013-2-1) levels were measured according to the manufacturer's instructions. Kidney tissues were fixed with 4 % paraformaldehyde, embedded in paraffin for HE and PAS staining, and then assessed under a microscope.
For immunohistochemical staining and immunofluorescence staining, paraffin-embedded kidneys were cut into 4 μm and 5 μm thick sections, deparaffinized with dimethylbenzene, and blocked with 10 % BSA. Then, the kidney sections were incubated with anti-F4/80 (1:100, Cat#: 28463-1-AP, Proteintech), anti-GPX4 (1:100, Abcam, ab125066), RAB7 (1:100, Cell Signal Technology, 9367S), PLIN2 (1; 100, Abcam, ab108323) and LC3B (1; 100, Cell Signal Technology, 83506) at 4 °C overnight. Histochemical images were captured under a microscope (Zeiss, Axio Vert A1).
GSH kits (A006-2-1, Nanjing Jiancheng) and MDA kits (A003-1-2, Nanjing Jiancheng) were used to detect the content of GSH and MDA in kidney or HK-2 cells according to the manufacturer's instructions.
Sample proteins were subjected to sodium dodecyl sulfate‒polyacrylamide gel electrophoresis in 10%–12.5 % gels, and the separated proteins were transferred to a 4.5 μm PVDF membrane. The PVDF membranes were blocked with 5 % skim milk for 1 h or Quick Block (P0220, Beyotime, China) for 20 min at room temperature. The quantification of the gray values of protein bands was dependent on ImageJ software. The relative expression of target proteins was normalized to ACTB/Tubulin.
Fresh kidney tissue was cut into 1 mm3 pieces and then fixed with 2 % glutaraldehyde. The fixation, dehydration, embedding, polymerization, and lead citrate staining were performed by the Core Facility of Basic Medical Sciences, Shanghai Jiao Tong University of Medicine. An H-7650 transmission electron microscope (Hitachi, H-7650) was used to detect 70-nm thick sections.
The frozen kidney sections were stained with Oil Red O stain (C0157S, Beyotime) for 30 min and then washed 3 times. Images were observed and captured under a microscope (Axio Zoom, V16, ZEISS).
The levels of TG and TC in serum and HK-2 cells were detected by kits (A110-1-1, A111-1-1, Nanjing Jiancheng) according to the manufacturer's instructions.
The viability of HK-2 cells was measured using a CCK-8 kit (Dojindo, CK04) according to the manufacturer's instructions. In brief, 10 μl of CCK-8 solution was added to HK-2 cells and incubated at 37 °C for 3 h. Absorbance at 450 nm was measured every 30 min using BioTak CytationTM3.
Lipid peroxidation was detected by BODIPY™ 581/591 C11 (10 μM, Thermo Fisher Scientific, D3861) added to HK-2 cells, which recognized oxidized and nonoxidized lipids. Free iron ions were stained with FerroOrange (1 μM, Dojindo, F374) to red, and the intensity of the fluorescence represents the level of label iron pools.
HK-2 cells were plated into 24-well plates on coverslips, followed by LPS for 24 h. For monitoring LDs and lysosomes, BODIPY (1 μM, D3922, Thermo Fisher Scientific) and LysoTracker (1 μM, 40739ES50, Yeasen Biotechnology, Shanghai) were added to HK-2 cells and then incubated for 30 min at 37 °C and 5 % CO2. Images of LDs and lysosomes were captured under an Olympus Laser Confocal Fluorescence Microscope (Olympus, Japan).
The sequences of small interfering RNA (siRNA) were as follows: RAB7-siRNA 5′-CCCAUUUGUUGUGUUGGGAAAttUUUCCCAACACAACAAAUGGGtt -3′. Transfection reagents (Lipofectamine 3000, L3000015, Thermo Fisher Scientific) were purchased from Thermo Fisher Scientific, and the experiments for siRNA were conducted according to the manufacturer's instructions.
IBM SPSS Statistics version 25 was used to analyze all statistical data. The qualitative data are presented as the mean ± standard error (SEM). A P value less than 0.05 was considered a significant difference, and a P value less than 0.01 was considered a significant difference.




留言 (0)