
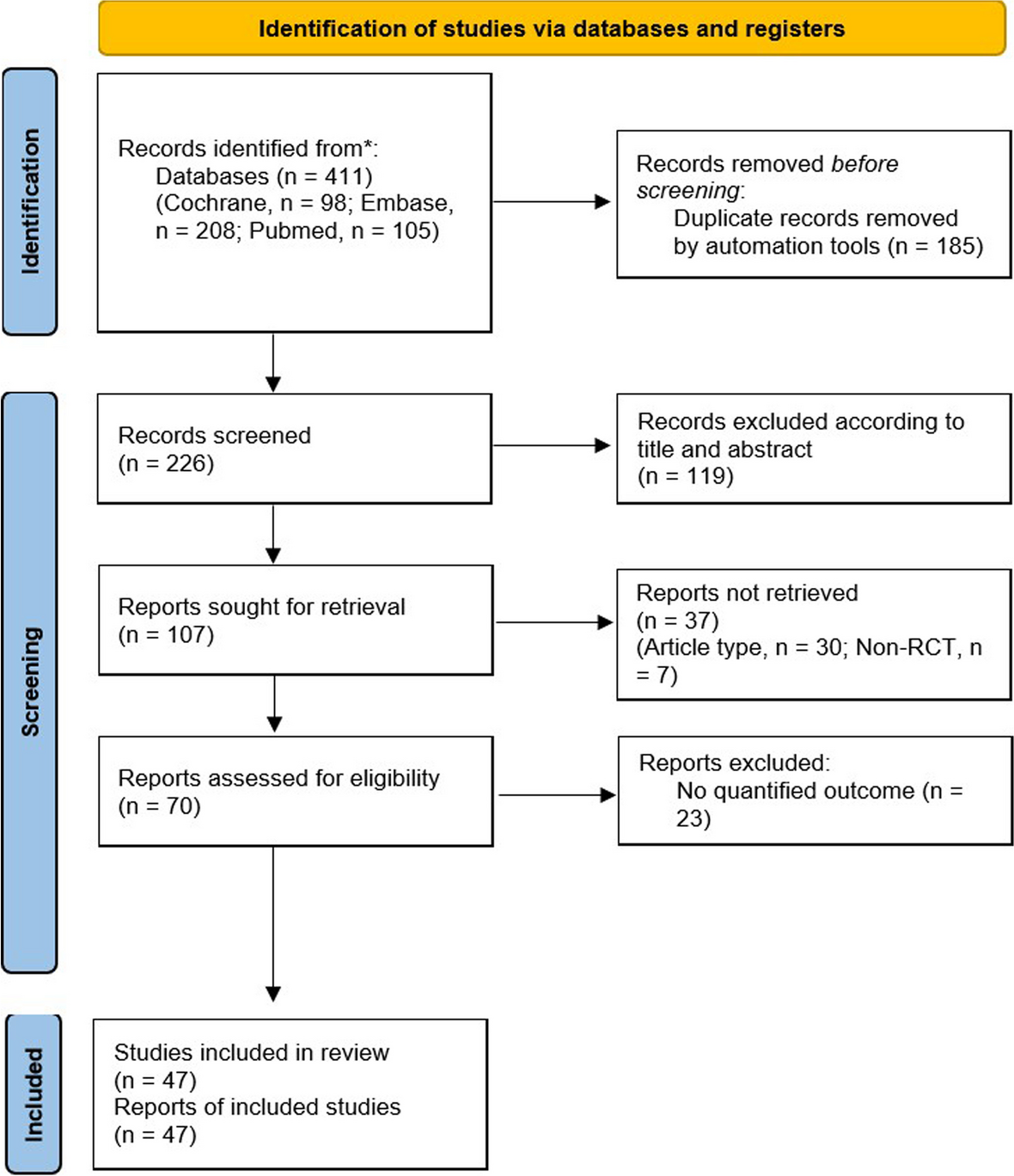
記住我




The LNCaP and PC3 cell lines treated with erastin were phenotypically different in the two cell groups (Fig. 1a). Erastin repressed the expression of the ferroptosis marker protein SLC7A11 in both PCa cell lines (Fig. 1b). Thus, we carried out cell survivability, MDA, Fe2+, GSH, and GSSG assays of erastin-induced PCa cells. After 48 h, erastin significantly inhibited the proliferation of LNCaP and PC3 cells in the CCK8 assay (p < 0.05; Fig. 1c). The results showed that MDA levels under erastin-induced conditions were notably increased in PC3 cells (p < 0.05; Fig. 1d). In contrast, erastin did not trigger a marked difference in MDA levels in LNCaP cells (p > 0.05; Fig. 1d). Fe2+ content, one of the key features of ferroptosis, was notably increased with erastin treatment in these two cell lines (p < 0.05; Fig. 1e). Given that GSH/GSSG regulates cellular redox homeostasis, the levels of GSH and GSSG were measured. The results showed that GSH levels in both cell lines were decreased following erastin treatment (p < 0.05; Fig. 1f). Meanwhile, the GSH/GSSG levels were also downregulated in these cell samples (p < 0.01; Fig. 1g).
Fig. 1
Erastin inhibits cell proliferation and induces ferroptosis in LNCaP and PC3 cells. (a) Phenotypic characterization of LNCaP and PC3 groups; “DMSO”, cells induced by DMSO as a negative control; “Erastin”, cells induced by erastin as an experimental group. (b) Expression of SLC7A11 was suppressed by erastin treatment in LNCaP and PC3 cells. (c) Proliferative levels of LNCaP and PC3 cells after erastin treatment. The relative MDA (d), Fe2+ (e), GSH (f), and GSH/GSSG (g) levels of LNCaP and PC3 cells treated with erastin were detected. Three independent experiments were performed with triplicate wells. Representative experiments and images are shown. Statistical significance was calculated using an independent-sample t-test. *p < 0.05, **p < 0.01, ***p < 0.001
Summary of RNA sequencing dataRNA sequencing was used to analyze the transcriptome profiling of PCa cells and erastin-induced PCa cells to investigate the molecular mechanisms by which the ferroptosis inducer erastin affects PCa cells and identify more FRGs. Using control cells and erastin-induced PCa cells, each of the four groups had three biological replicates, and 12 sequencing samples were prepared. Three replicates in four groups were consistent with the principal component analysis (Fig. 2a). Based on the correlation analysis results (Fig. 2b), the correlation coefficient of the samples was between 0.8 and 1, which showed good biological repeatability in these sequencing samples.
Erastin-induced DEGs analysisIn the RNA-seq assay with padj ≤ 0.05 and |log2(foldchange)|≥2 as screening conditions, LNCaP_5_0_Era identified a total of 942 DEGs compared with LNCaP, including 447 upregulated genes and 495 downregulated genes (Fig. 2c–d). Then, a total of 5,625 DEGs were screened and recognized between PC3_5_0_Era and PC3, of which 3,704 were upregulated and 1,921 were downregulated (Fig. 2e–f). A total of 295 overlapping DEGs in the two compared groups were screened and recognized via a Venn diagram (Fig. 2g).
Fig. 2
DEGs identification in LNCaP and PC3 cells after erastin exposure. (a) Principal component analysis (PCA) of RNA-seq data. (b) The correlation analysis results were consistent with PCA. (c) Volcano plot analysis identifies the DEGs of erastin-induced LNCaP groups; red and green dots represent 447 and 495 upregulated and downregulated genes, respectively. (d) Heatmap of LNCaP groups after erastin exposure. (e) Volcano plot analysis identifies the DEGs of PC3 groups; red and green dots represent 3,704 and 1,921 upregulated and downregulated genes, respectively. (f) Heatmap for PC3 groups after erastin exposure. (g) Venn diagram of erastin-induced DEGs
Functional enrichment of erastin-induced genes and validationGO, KEGG, and REACTOME functional enrichment analyses were performed to analyze 295 overlapping DEGs to further explore the potential biological behavior of erastin-induced changes in PCa cells. As exhibited by the GO analysis results, the erastin-induced DEGs were primarily related to DNA replication, DNA-dependent DNA replication, and DNA replication initiation for biological processes (BPs), condensed chromosome, chromosomal region, condensed chromosome, centromeric region for molecular functions (MFs), and catalytic activity acting on DNA, DNA helicase activity, and helicase activity for cellular components (CCs) (Fig. 3a). In contrast, KEGG analysis indicated that these DEGs mapped to DNA replication, cell cycle, and homologous recombination pathways (Fig. 3b). Furthermore, REACTOME analysis revealed that these DEGs were associated with DNA strand elongation, cell cycle checkpoints, activation of the prereplicative complex, and so on (Fig. 3c). To validate the functional enrichment analyses, the flow cytometry was employed to measure the cell cycle distribution. According to the flow cytometry modulation results, the percentage of PCa cells in G1 phase was considerably greater in the erastin-treated group than in the control group (p < 0.05; Fig. 3d).
Fig. 3
Enrichment analysis of the DEGs in erastin-induced LNCaP and PC3 cells. (a) GO functional analysis. BP, CC, and MF represent biological processes, cellular components, and molecular functions, respectively. (b) KEGG pathway analysis. (c) RECTOME functional analysis. (d) Flow cytometry detection of the cell cycle distribution of PCa cells after treatment with erastin. The results are expressed as a peak diagram, and the distributions of cells in the G1, S, and G2 phases were calculated. Statistical significance was calculated using an independent-sample t-test. *p < 0.05, **p < 0.01
Pathways and module analysis of two PCa cellsAn MCODE plug-in was carried out to construct modules representing the crucial clusters to access which related pathways contributed more weight to the ferroptotic process of PCa cells (Supplementary Fig. 1). Among the LNCaP group, the DEGs in module 1 were selected to conduct the next analysis, and they were found to be involved in several prominent signaling pathways (Supplementary Fig. 1a–b). For the PC3 group, enriched pathways, including nephron morphogenesis, positive regulation of blood vessel endothelial cell migration, positive regulation of bone mineralization, and regulation of steroid metabolic process, were detected in module 1 of PC3 DEGs (Supplementary Fig. 1c–d).
GSEA analysis of erastin-induced genesHerein, we then performed GSEA analysis between the erastin treatment and control groups across the four cell lines to reveal more gene set enrichment information that involves the erastin-induced changes in PCa cells. We found that most of those pathways in GSEA were also found in the GO, KEGG, and Reactome functional enrichment analyses, which supports and validates the previous results. However, it was not unexpected to find that erastin-induced PCa DEGs were involved in the regulation of cell death, cellular response to reactive oxygen species, fatty acid biosynthetic process, activation of the MAPK activity pathway, intrinsic apoptotic signaling pathway in response to DNA damage by the P53 class mediator and cellular amino acid metabolic process in LNCaP (Fig. 4a) and PC3 cells (Fig. 4b). Moreover, we further focused on several pathways expected to be induced only in the PC3 group, including regulation of I-kappab kinase NF-kappab signaling, regulation of the JNK cascade, regulation of the ERK1 and ERK2 cascades, negative regulation of the ERBB signaling pathway, negative regulation of Ras protein signal transduction, and regulation of the JAK-STAT cascade (Fig. 4c).
Fig. 4
GSEA analysis based on the RNA-seq datasets. (a) GSEA analysis shows enrichment of the response genes in LNCaP group after erastin exposure. (b) GSEA analysis results of the enrichment of response genes in the PC3 group with erastin treatment. (c) GSEA analysis of enrichment of the response genes, which only appeared in the erastin-induced PC3 group. NES, normalized enrichment score; nominal p-value, and FDR, false discovery rate, and q-value were determined by GSEA software
Erastin-induced hub genes and their TFsThe cytoHubba plug-in was selected to calculate the hub genes of 295 overlapping DEGs to find more potential and pivotal genes related to ferroptosis in PCa cells. Of these, 10 top hub genes (including CLU, IL1B, MET, NRXN3, PLXNA4, GAD1, UNC5B, SLC7A5, DAPK1, and TMEFF2) were identified in order of degrees (Fig. 5a). Details and descriptions of the 10 hub genes are summarized in Table 1.Given that TFs had a critical influence on gene expression levels, the 10 top hub genes were uploaded to the NetworkAnalyst database for TF prediction. The regulatory network of the top TFs (including SP9, DLX2, ARX, PEG3, CSRNP3, ZNF697, INSM2, STAT3, NR2F1, FOXL1, E2F1, and NFIC) of the hub genes was obtained and presented (Fig. 5b). Moreover, the expression levels of these TFs are shown in Fig. 5c. Among them, PEG3, CSRNP3, ZNF697, NR2F1, DLX2, and INSM2 were highly expressed, whereas SP9, ARX, E2F1, FOXL1, STAT3, and NFIC were expressed at low levels in PCa cells after erastin exposure. The expression levels of a few hub genes and TFs (including CLU, IL1B, UNC5B, PEG3, DLX2, SP9, STAT3, E2F1, and NFIC) were validated by qPCR (Fig. 5d). Moreover, we analyzed their expression patterns in the HPA and GEPIA databases to investigate whether these hub genes were clinically associated with PCa. In the HPA database, the expression of TMEFF2 in PCa tissue was higher than that in normal tissue, and the expression levels of CLU, NRXN3, and UNC5B were lower in PCa tissue by immunohistochemistry (Fig. 5e). Consistently, TMEFF2 mRNA levels were higher in PCa tissue (p < 0.05), and CLU, NRXN3, and UNC5B were downregulated (p < 0.05) in PCa tissue in the GEPIA database (Fig. 5f).
Table 1 Top 10 hub genes of erastin-induced LNCaP and PC3 cellsFig. 5
Prediction and validation the expression of TFs targeting hub genes and the clinical characteristics of the essential hub genes. (a) Top 10 hub DEGs of the LNCaP and PC3 cell groups with erastin treatment. (b) Interaction network of hub genes and their TFs. (c) The heatmap of the expression of the TFs was revealed by RNA-seq. (d) Expression of hub DEGs and TFs after erastin treatment were measured by qPCR, n = 3. The expression levels of TMEFF2, CLU, NRXN3, and UNC5B in normal tissues and prostate cancer tissue from the HPA database (e) and GEPIA database (f). *p < 0.05, *p < 0.01, ***p < 0.001
Prediction and validation of FRGsThe relationships between hub gene expression levels and the ferroptosis markers SLC7A11 and GPX4 in the GEPIA database were analyzed to investigate whether these 4 hub genes were potentially involved in ferroptosis. As expected, the expression patterns of TMEFF2, CLU, NRXN3, and UNC5B were positively correlated with SLC7A11 and GPX4 (Fig. 6a). Gene interaction networks were established to clarify the possible direct relationship between hub genes and ferroptosis markers and identify their potential associations. TMEFF2, SLC7A11, and GPX4 showed a complex PPI network with physical interactions, co-expression, prediction, co-localization, genetic interactions, pathways, and shared protein domains of 77.64%, 8.01%, 5.37%, 3.63%, 2.87%, 1.88%, and 0.60%, respectively (Fig. 6b). Among these networks, there were direct genetic interactions between TMEFF2 and SLC7A11. Moreover, TMEFF2 showed direct genetic interactions with ALOX5, and ALOX5 showed physical interactions with GPX4. However, CLU, NRXN3, and UNC5B failed to display such a close relationship with GPX4 compared with TMEFF2 (data not shown). Therefore, TMEFF was selected as an interesting ferroptosis-related gene for further experiments.
In addition, we constructed LNCaP, VCaP, PC3 and C4-2 cells with TMEFF2 downregulation to test whether TMEFF2 can be an FRG in the experiment and then detected their cell survivability and Fe2+ concentration. Androgen-dependent LNCaP and VCaP cell lines with knockdown of TMEFF2 exhibited lower expression of SLC7A11, while downregulation of TMEFF2 did not significantly affect the expression level of SLC7A11 in androgen-independent PC3 and C4-2 cell lines (Fig. 6c). After 48 h, the downregulation of TMEFF2 significantly reduced the proliferative ability of LNCaP and VCaP cells (p < 0.05; Fig. 6d). After TMEFF2 was knocked down, the level of Fe2+ in LNCaP and VCaP cells was markedly increased (p < 0.05; Fig. 6e). In contrast, there was no significant difference between PC3 and C4-2 cells with erastin treatment and the control group (Fig. 6e). To demonstrate the androgen-regulated property of TMEFF2, enzalutamide was selected to suppress the AR expression in LNCaP and VCaP cells. These cells were exposed to 10 µM enzalutamide for 48 h, which inhibited AR expression but not the expression of SLC7A11 (Fig. 6f). LNCaP and VCaP cells with knockdown of TMEFF2 and enzalutamide treatment, the expression of SLC7A11 was not significantly different compared to cells with down-regulation of TMEFF2 (Fig. 6f). Then, GSE35988 was utilized to confirm TMEFF2 expression in clinical prostate tissue. Compared with normal tissues, the expression pattern of TMEFF2 was markedly increased in localized prostate cancer. However, the TMEFF2 expression levels did not significantly differ between normal tissues and CRPC tissues (Fig. 6g).
Fig. 6
Correlation analysis and validation of FRGs. (a) Correlation analysis between four DEGs and SLC7A11 and GPX4. (b) Gene co-expression network analysis of TMEFF2, SLC7A11, and GPX4. (c) The expression of SLC7A11 was suppressed by downregulating TMEFF2 in LNCaP and VCaP cells but not in PC3 and C4-2 cells. (d) The cell growth of LNCaP, VCaP, PC3 and C4-2 cells with knockdown of TMEFF2 was examined via a CCK8 assay. Three independent experiments were performed with triplicate wells. Representative experiments and images are shown.(e) The relative Fe2+ concentration of LNCaP, VCaP, PC3 and C4-2 cells with downregulation of TMEFF2 was detected. (f) After knockdown of TMEFF2 and enzalutamide treatment, the expression of SLC7A11, TMEFF2, and androgen receptor (AR) were detected by western blot. (g) The mRNA expression of TMEFF2 in normal tissue and prostate cancer tissue (including localized prostate tissue and CRPC tissue) was obtained from the GSE35988 dataset in the GEO database. The statistical significance of differences was calculated using an independent-sample t test. *p < 0.05, **p < 0.01
留言 (0)