
記住我
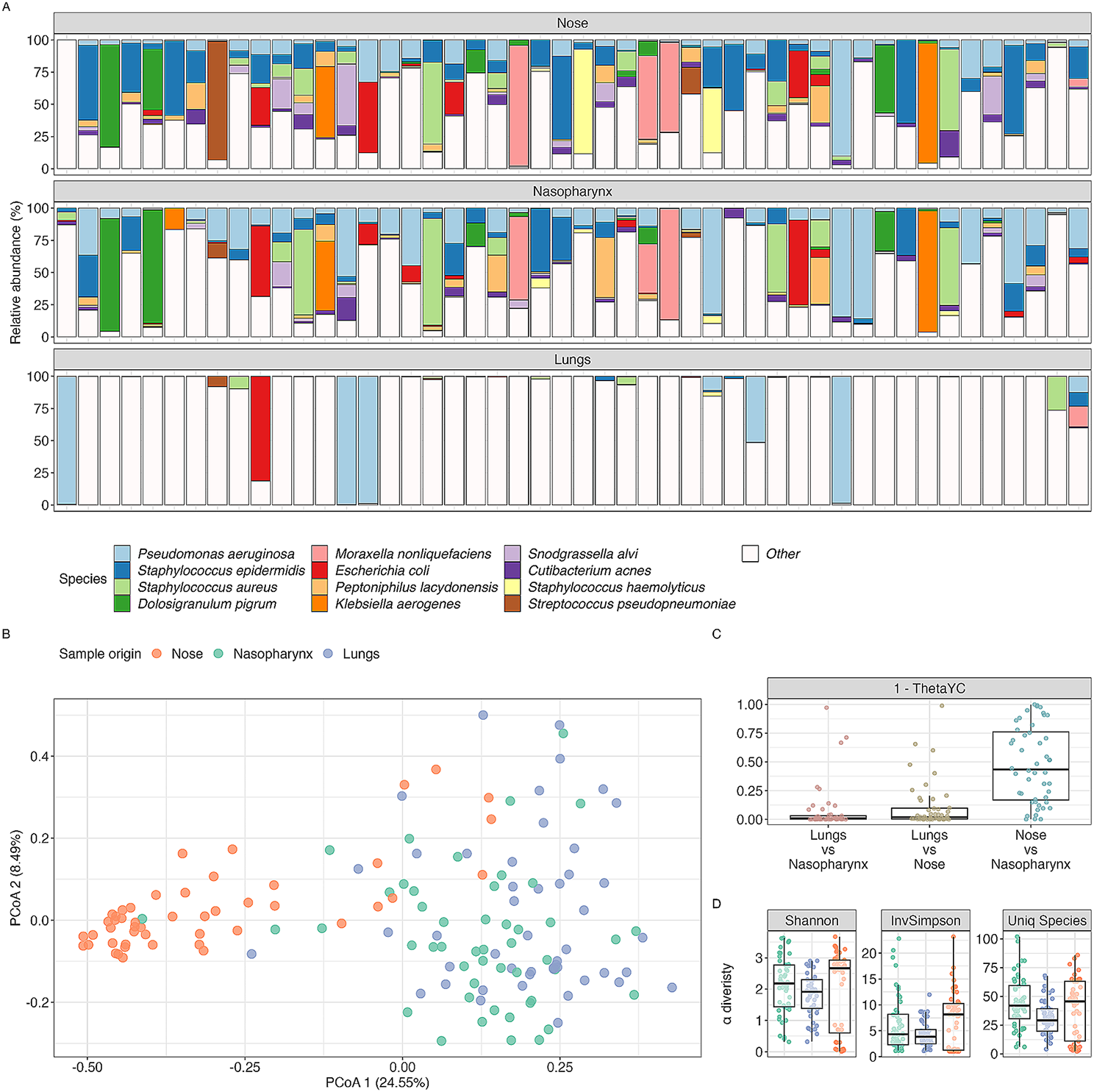



A total of 23 distinct strains were isolated from SOM based on the isolation process. Among these, nine strains demonstrated notable antipathogenic properties, effectively inhibiting the growth of the listed human enteropathogens, as shown in Supplementary Figure S3.
Antipathogenic Activities (Agar Well Diffusion Assay)Out of the 23 strains isolated from SOM, nine isolates exhibited notable inhibitory activities against common human enteropathogens. The results of the inhibitory activities of these nine strains are presented in Table 2. The raw data on the diameters of the inhibition zones are provided in Supplementary Table S1.
Table 2 Isolated strains’ inhibition to human enteropathogensWithin this group of nine strains, eight were identified via 16S rRNA sequencing as members of the B. subtilis species, with the remaining one strain classified as Weissella paramesenteroides. Among these strains, B. subtilis SOM8 displayed the most promising inhibitory activities against all five selected human enteropathogens. Notably, B. subtilis SOM8 exhibited remarkable inhibitory effects on the growth of common foodborne pathogens, including V. parahaemolyticus and B. cereus, which can cause diarrhoeal diseases, as well as the virulent serotype E. coli O157:H7, responsible for diarrhoea and associated complications. Furthermore, B. subtilis SOM8 exhibited inhibitory effects on not only Gram-positive pathogens (S. aureus, and B. cereus) but also Gram-negative pathogens (E. coli, S. enterica, and V. parahaemolyticus). Consequently, B. subtilis SOM8 was selected for further comprehensive phenotypic and genotypic screening.
Taxonomic Information of B. subtilis SOM8Phylogenetic analysis was employed to discover the relationship between B. subtilis SOM8 and several closely related strains. A phylogenetic tree of B. subtilis SOM8 was constructed using TYGS, as illustrated in Fig. 1, the raw data of TYGS results are shown in Supplementary Table S2 and S3. B. subtilis SOM8 was found to share close phylogenetic proximity with well-known wild-type strains, notably B. subtilis NCIB 3610 and B. subtilis ATCC 6051. Moreover, in alignment with the BLAST results in prior research, B. subtilis SOM8 also demonstrated a close taxonomic alignment with B. subtilis subsp. subtilis 168, as well as a commercially available probiotic strain, B. subtilis MB40 [10]. These findings underscore the substantial potential of isolated B. subtilis SOM8 for application as a probiotic.
Fig. 1
Phylogenetic tree of B. subtilis SOM8 with similar Bacillus strains using TYGS database
Acid and bile toleranceThe results depicting survival of B. subtilis SOM8 after exposure to acid (pH 2, 3, and 4) and varying concentrations of ox-bile salts (0.5%, 1.0%, 1.5% w/v), as well as mixed acid and bile salt conditions for 2 h, are presented in Figs. 2 and 3. Notably, B. subtilis SOM8 exhibited susceptibility to low pH conditions (pH 2 and pH 3), resulting in a reduction of 4 to 5 Log10CFU/mL. However, under pH 4, the reduction in Log10CFU/mL was less than 1, indicating a good survivability to acidic environments when the pH exceeded 4. In the context of bile salts, B. subtilis SOM8 demonstrated Log10CFU/mL reductions ranging between 2 and 3 across various concentrations, indicating its great tolerance to bile salts. Furthermore, it is noteworthy that under both pH 4 and bile salt conditions, the final Log10CFU/mL count for B. subtilis SOM8 remained consistently above 6. This observation suggests the considerable potential of B. subtilis SOM8 to establish a colony within the human GIT, thereby contributing to its functional role [52]. In addition, when B. subtilis SOM8 was exposed to mixed acid and bile salt conditions, the survival pattern was closely related to that observed under solely acidic stress conditions, emphasizing the dominant role of acid in affecting the survivability of B. subtilis SOM8.
Fig. 2
B. subtilis SOM8 tolerance to acid and bile salt. Log10CFU/mL of B. subtilis SOM8 was measured before and after exposure to acid or ox-bile salts for 2 h, respectively
Fig. 3
B. subtilis SOM8 tolerance to mixed acid and bile salts conditions. Log10CFU/mL of B. subtilis SOM8 was measured before and after exposure to acid with ox-bile salts for 2 h, respectively
SGF/SIF ToleranceThe susceptibility of B. subtilis SOM8 to SGF and SIF was assessed to simulate the gastrointestinal conditions, with the results presented in Fig. 4. B. subtilis SOM8 demonstrated robust survival in SIF, maintaining a Log10CFU/mL count exceeding 8 after 2 h. Conversely, the strain exhibited relative susceptibility to SGF due to its low acidic environment (pH 2) and the presence of porcine pepsin, resulting in a reduction of Log10CFU/mL between 4 and 5. Nonetheless, the survivability remained at approximately 50%, signifying a better tolerance to both SGF and SIF than mostly applied commercial probiotic strain LGG [53]. Therefore, B. subtilis SOM8 exhibits substantial potential for applications as probiotics, with the possibility of encapsulation to enhance its survivability under human GIT.
Fig. 4
B. subtilis SOM8 tolerance to SGF and SIF, respectively
Heat stabilityThe outcomes of this investigation are presented in Fig. 5. Evidently, the figure depicts that the amount of the bacteria remained constant under 40 °C, with Log10CFU/mL reduction smaller than 0.1. As the temperature elevated to 60 °C, a minor decline in the Log10CFU/mL from 8.3 to 7.8 was observed, with survivability retained at more than 90%. However, a substantial decline was observed as the temperature reached 80 °C, leading to a reduction in the Log10CFU/mL to 4.7. Notably, the exposure of the bacteria to 100 °C for the same duration resulted in complete cell inactivation.
Fig. 5
B. subtilis SOM8 tolerance to elevated temperatures from 40 °C to 100 °C for 30 min
Antioxidant Activity (DPPH Scavenging Assay)The antioxidant activities of the cell culture, supernatant, and PBS-resuspended cells of B. subtilis SOM8 are illustrated in Figs. 6 and 7, respectively. In Fig. 6, it reveals that B. subtilis SOM8 in TS broth, exhibited a remarkable antioxidant activity, evidenced by an approximate 40% DPPH scavenging, similar to that of L. plantarum. Contrarily, B. subtilis SOM8 in MRS broth resulted in low antioxidant activity, with DPPH scavenging ranging between 10 and 15%. Notably, all PBS-Resuspended cells including dissoluble metabolites displayed relatively lower antioxidant activity, while the original cell culture and the supernatant exhibited comparatively high antioxidant activity. This discrepancy could be attributed to the fact that the antioxidant activity primarily originates from secondary metabolites present in the supernatant, such as the exopolysaccharide (EPS) and organic acids produced by the cells. When B. subtilis SOM8 is introduced into MRS, a broth with relatively lower pH that is usually used for growing lactic acid bacteria, it appears to augment biofilm production [54]. Such a strategy is usually employed by microbes to overcome harsh environments like low pH, rather than synthesizing secondary metabolites for antioxidant activity. In Fig. 7, the data illustrates a progressive enhancement in antioxidant activity, reflected by the DPPH scavenging increasing from 40 to 60%, as supplementary sucrose is incrementally introduced into the TS broth, up to a concentration of 150 g/L. Therefore, there exists a saturation point, beyond which adding more sucrose will have no effect on antioxidant activity. The results also proved the dominant role of sucrose instead of monosaccharide in producing EPS [55].
Fig. 6
Antioxidant activity of B. subtilis SOM8 in TS, MRS broth, respectively
Fig. 7
Antioxidant activity of B. subtilis SOM8 in TS broth with different concentrations of supplemented sucrose
BSH activityThe BSH activity of isolated B. subtilis strains SOM 1–8 were shown in Supplementary Figure S4. Evidently, all eight strains of isolated B. subtilis exhibited BSH activities, as signified by the white precipitation surrounding the colonies. In contrast to their growth on standard TS agar plates devoid of TDC supplementation, the morphology is notably distinct. The formation of this white precipitate around the colonies underscores the enzymatic deconjugation of bile salts to primary bile salts by these strains [56].
Haemolytic activityThe haemolytic activity of isolated B. subtilis SOM8 and wild type strain B. subtilis ATCC 6051 was shown in Supplementary Figure S5. Both isolated B. subtilis SOM8 and wild-type B. subtilis ATCC 6051 exhibited α-haemolytic activity, characterized by partial or green haemolysis linked to the reduction of red cell haemoglobin. This phenomenon is attributed to the production of hydrogen peroxide by the bacterium such as S. pneumoniae, causing oxidation of iron in haemoglobin and resulting in the formation of the green oxidized derivative, methaemoglobin [57].
Cell cytotoxicity using Caco-2 cells (CCK-8 Assay)The results depicting cell cytotoxicity resulting from exposure to the cell-free filtrate and lyophilized cell-free filtrate of B. subtilis SOM8 and B. subtilis ATCC 6051 are illustrated in Figs. 8 and 9, respectively.
Fig. 8
The effects of B. subtilis SOM8 and B. subtilis ATCC 6051 cell-free filtrate on viability of Caco-2 cells
Fig. 9
The effects of B. subtilis SOM8 and B. subtilis ATCC 6051 freeze-dried cell-free filtrate on viability of Caco-2 cells
Notably, both the cell-free filtrate and freeze-dried filtrate of B. subtilis SOM8 and B. subtilis ATCC 6051 exhibited low cytotoxicity. At concentrations of 5 µL/mL or 10 mg/mL, an observable trend indicated that the filtrates demonstrated to inhibit the proliferation of Caco-2 cells. This phenomenon might be attributed to the presence of bioactive compounds and hydrogen peroxide produced by both strains of B. subtilis. Furthermore, the substitution of the fermented solution for DMEM was identified as a contributing factor to the observed inhibition, as evidenced by a relatively higher inhibitory effect for the cell-free filtrate compared to the freeze-dried filtrate. In summary, considering the low cytotoxicity towards Caco-2 cells, both B. subtilis SOM8 and B. subtilis ATCC 6051 are viable candidates for probiotic applications, with B. subtilis SOM8 exhibiting comparatively better performance.
Adhesion capacity assay using caco-2 cellsThe adhesion capabilities of B. subtilis SOM8, B. subtilis ATCC 6051, and LGG to Caco-2 cells are graphically represented in Fig. 10. Notably, adhesion capacity exhibits no apparent correlation with the initial seeding concentration. Interestingly, all three strains demonstrated optimal adhesion at an initial seeding concentration of 107 CFU/mL, with B. subtilis SOM8 owning an approximately 70% adhesion rate, significantly higher than both B. subtilis ATCC 6051 of 20% and LGG of 8%. This heightened adhesion exhibited by B. subtilis SOM8 is possible to be attributable to its augmented biofilm-producing and intrinsic adhesion properties. Importantly, it is observed that B. subtilis SOM8 consistently meets the required criteria for probiotic efficacy, maintaining adhesion values surpassing 106 CFU/mL across varying seeding concentrations. This adherence threshold, as established guidelines [52], indicated B. subtilis SOM8's fulfilment of the necessary criteria for optimal probiotic functionality under diverse seeding conditions.
Fig. 10
Adhesion capacity of B. subtilis SOM8, B. subtilis ATCC 6051 and LGG to Caco-2 cells under different initial seeding concentrations
WGS of B. subtilis SOM8 for Genotypic CharacterizationVirulence factors identificationThe prediction of virulence factors within B. subtilis SOM8 was facilitated through the VFDB database. As illustrated in Table 3, the genome of B. subtilis SOM8 revealed a total of 10 matches with VF-associated protein. B. subtilis SOM8 was observed to lack the B. cereus cereulide gene cluster (cesABCHPT) and the enzyme genes encoded by pathogenic Bacillus species. The virulence factor of B. subtilis SOM8 involves toxins, immune evasion, and iron acquisition. Among the identified virulence factors, the iron acquisition, related genes dhbABCDE were identified within the genome of B. subtilis SOM8. However, these dhbA-E genes are a common genetic component in B. subtilis subsp. subtilis 168, a strain extensively utilised in industrial applications. Notably, B. subtilis SOM8 was also predicted to encode for capABCD genes, implicated in polyglutamate synthesis and transport. However, the capE gene, present in B. anthracis and B. cereus biovar anthracis, is absent. The final identified virulence factor is toxins. The B. subtilis SOM8 was predicted to encode a haemolysin, putative membrane hydrolase (hlyIII), based on genetic information. Notably, comparable haemolytic activity has been detected across several Bacillus strains, including those utilised as commercial probiotics [58]. Furthermore, the likelihood of an orally administered probiotic translocating through the intestinal barrier into the bloodstream remains limited and has been reported only at minimal frequencies in hospitalized patients [59].
Table 3 Summary of isolated B. subtilis SOM8 genome for virulence factor prediction using VFDB databaseIn the comparative analysis of VFs between isolated B. subtilis SOM8 and B. subtilis ATCC 6051 a striking similarity in VFs was observed. However, it was found that B. subtilis SOM8 lacks the bpsC gene responsible for B. cereus EPS production that exists in B. subtilis ATCC 6051, a specific immune evasion VF present in B. cereus. This absence of the bpsC gene in B. subtilis SOM8 highlights its enhanced safety profile compared to B. subtilis ATCC 6051.
Antibiotic resistance genes identificationThe evaluation of antibiotic resistance genes within B. subtilis SOM8 was undertaken utilizing the CARD. Out of a total of 274 hits, 10 hits exhibited a minimum identity of 95% and were subsequently categorized as strict matches, as shown in Supplementary Table S4. Notably, the remaining hits displaying identity levels below 80% were not taken into consideration. For instance, with an identity of 98.59% to the aadK gene, the isolated B. subtilis SOM8 is predicted to exhibit resistance against streptomycin. Additionally, its resistance to macrolides spiramycin and telithromycin can be attributed to a 98.35% identity with the gene mphK, which encodes a macrolide phosphotransferase. In summary, B. subtilis SOM8 was predicted to harbour 10 antibiotic resistance genes, conferring potential resistance against a diverse spectrum of antibiotics. These encompass peptides, fluoroquinolones, aminoglycosides, tetracyclines, phenicols, lincosamides, nucleosides, macrolides, streptogramins antibiotics as well as disinfecting agents and antiseptics. Nevertheless, it is imperative to acknowledge that gene prediction does not necessarily imply gene expression. To address this critical aspect, MIC assessments against a spectrum of medically significant antibiotics were conducted.
Plasmid and MGEs IdentificationDrawing from the outcomes obtained through the PlasmidFinder 2.1 [40] and MobileElementFinder [41], it is evident that B. subtilis SOM8 lacks plasmid genes and any MGEs. The inference can be drawn that this strain may not possess the capability to transfer potential antibiotic resistance genes to other bacterial entities. It is necessary to acknowledge, however, that these conclusions stem from BLAST-based assessments and genetic data. For a more comprehensive understanding, the need for in vivo investigations or subsequent clinical trials is still necessary, especially in anticipation of the eventual integration of this strain into both industrial and medical applications.
MIC Evaluation of B. subtilis SOM8The sensitivity of B. subtilis SOM8 to eight medically prescribed antibiotics was tested following CMSI and EFSA MIC standard, the results were shown in Table 4. The investigation revealed that isolated B. subtilis SOM8 displayed susceptibility to seven out of eight common antibiotics, including one glycopeptide, two aminoglycosides, two macrolides, one tetracycline, and one phenicol antibiotic, in accordance with EFSA standards. The MIC of B. subtilis SOM8 to streptomycin was approximately 128 μg/mL, notably exceeding the EFSA threshold. However, it is essential to notify that resistance to streptomycin is generally regarded as an intrinsic property of Bacillus species that contain the putative aadK genes. Moreover, there is no supporting evidence indicating the potential horizontal transfer of such genes to other bacterial strains [60].
Table 4 MIC results of isolated B. subtilis SOM8 against eight common antibioticsSecondary Metabolites (antiSMASH, BAGEL4) PredictionThe assessment of secondary metabolites, including bacteriocins, synthesized by the isolated B. subtilis SOM8 was conducted using the antiSMASH and BAGEL4 databases, as detailed in Table 5. B. subtilis SOM8 was predicted to yield six distinct secondary metabolites, encompassing both Ribosomally Synthesized and Post-Translationally Modified Peptides (RiPPs) and Non-Ribosomal Peptide Synthases (NRPS). These include fengycin, bacillaene, subtilosin, bacilysin, bacillibactin, and lichendicin.
Table 5 Summary of predicted secondary metabolites produced by isolated B. subtilis SOM8In contrast to primary metabolites, these secondary metabolites represent non-essential, small organic molecules that can potentially confer evolutionary advantages over time, such as enhancing survival in competition with other organisms. B. subtilis SOM8 is predicted to engage in the synthesis of diverse bioactive molecules, notably encompassing various antibiotics with considerable potential for applications. These findings underscore the SOM’s capacity to produce an array of compounds with potential therapeutic applications.
Comparison with Wild Type Strain B. subtilis ATCC 6051The growth patterns of B. subtilis SOM8 and the wild-type strain B. subtilis ATCC 6051 were compared under both aerobic and anaerobic conditions, as depicted in Supplementary Figure S6. Additionally, their respective antipathogenic activities against the specified human enteropathogens were tested under both aerobic and anaerobic conditions, as presented in Supplementary Figure S7. The results underscored B. subtilis SOM8's superior growth performance under both aerobic and anaerobic conditions. Notably, the B. subtilis SOM8 colonies exhibited enhanced dimensions and were surrounded by a more substantial excretion of biofilm, a matrix of extracellular substances known to create a favourable microenvironment for bacterial proliferation, particularly in challenging conditions.
The outcomes from Supplementary Figure S7 emphasize that both isolated B. subtilis SOM8 and B. subtilis ATCC 6051 own a wide spectrum of antipathogenic activities against several pathogens. However, it is noteworthy that B. subtilis SOM8 outperforms B. subtilis ATCC 6051 under anaerobic conditions. Specifically, when cultivated under anaerobic conditions (represented by Number 5 and 6), B. subtilis ATCC 6051 exhibits a loss of antipathogenic efficacy against V. parahaemolyticus and S. aureus, whereas B. subtilis SOM8 sustains its robust inhibition of pathogen growth, as evidenced by the inhibition zones.
留言 (0)