
記住我

The threat of cancer to human health continues, highlighting the vital importance of early detection, diagnosis and treatment in tumor management. However, the challenge in early diagnosis and treatment arises from the inconspicuous symptoms and lack of specificity in early-stage tumors. The current surgery, chemoradiotherapy, targeted therapy, and combined treatment in a variety of ways have greatly improved the therapeutic effect of tumors, however, due to the severe side effects and drug resistance, their efficacy remains unsatisfactory (1, 2). Hence, there is an imperative to innovate and develop novel treatments that can overcome these limitations. The advent of immunotherapy has revolutionized cancer treatment, owing much of its success to the breakthroughs in immune checkpoint blockade.
Programmed cell death-1(PD-1) is a crucial immunosuppressive molecule with two known ligands, programmed cell death-ligand 1(PD-L1) and PD-L2. The interaction between PD-1 and its ligands results in two opposing effects. One effect is the inhibition of T-cell immunity and the reduction of unnecessary immune responses, contributing to the prevention of autoimmune diseases. Another is the inhibition of the immune system’s ability to monitor and clear tumor cells, thereby allowing the occurrence and development of tumors (3). At present, many studies have been carried out on the PD-1/PD-L1 axis. Certain drugs, including Nivolumab and Pembrolizumab, are employed in the treatment of malignant tumors. However, it has been observed during treatment that not all PD-L1-positive patients exhibit a discernible response to anti-PD-1/PD-L1 axis treatment. Conversely, some PD-L1-negative patients show a positive response to the anti-PD-1/PD-L1 axis treatment, indicating that there may be other molecules or receptors that interact with PD-1 (4).
As the second known ligand of PD-1, whether PD-L2 can be used as another target for tumor therapy has attracted the attention of many researchers. As research progresses, potential interactions and relationships between PD-L1 and PD-L2 emerge. Simultaneous or separate expression of these two ligands can yield varied outcomes in terms of tumor development and prognosis. In a study involving 172 patients with head and neck tumors treated with pembrolizumab, the overall response rate (ORR) was two times higher in both PD-L1 and PD-L2 positive patients than in patients positive for PD-L1 alone. When PD-L2 expression was confined to tumor cells, the ORR was 26.5% in PD-L2 positive patients compared to 16.7% in PD-L2 negative patients (4). These findings indicate a potential relationship between PD-L1 and PD-L2, highlighting the need for investigating anti-PD-1/PD-L2 axis therapy. This may enhance the therapeutic efficacy of tumors when combined with anti-PD-1/PD-L1 axis therapy.
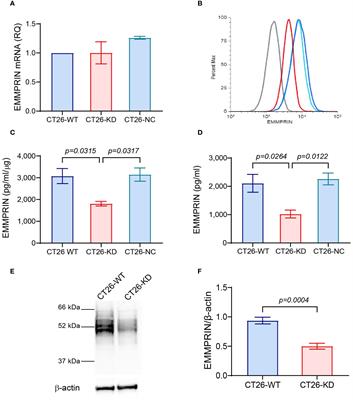
The binding affinity of PD-1 with PD-L1 and PD-1 with PD-L2 is quite different. While both ligands exhibit binding affinities to PD-1, structural analyses reveal significant differences in their interaction mechanisms. The binding of PD-1 and PD-L1 induces complex molecular configuration changes(Figure 1A), whereas the interaction between PD-1 and PD-L2 is simpler (5). Surface plasmon resonance analyses highlight that PD-L2 demonstrates 2 to 6 times higher affinity for PD-1 compared to PD-L1 (6), and in some cases, the affinity is reported to be 30 times higher (7). PD-L1 and PD-L2 not only bind to PD-1, but also have their own second binding sites. PD-L1 binds to CD80 (8) while PD-L2 binds to RGMb (9). CD80 functions as a co-stimulatory factor during T lymphocyte activation when CD86 activates, playing a pivotal role in autoimmune surveillance, humoral immune response, and transplantation reactions. The interaction between CD80 and PD-L1 results in heterodimerization, thereby impeding the interaction between PD-L1 and PD-1 (Figure 1B). However, the interaction of PD-L2 with PD-1 is not negatively regulated by the level of CD80 (10). Therefore, this also explains to some extent that anti-PD-L1 therapy is not effective for all PD-L1 positive patients, which further highlights the necessity and importance of studying PD-L2 and anti-PD-L2 drugs. This heightened affinity has implications for the development of small-molecule compound drugs and the PD-1/PD-L2 axis.
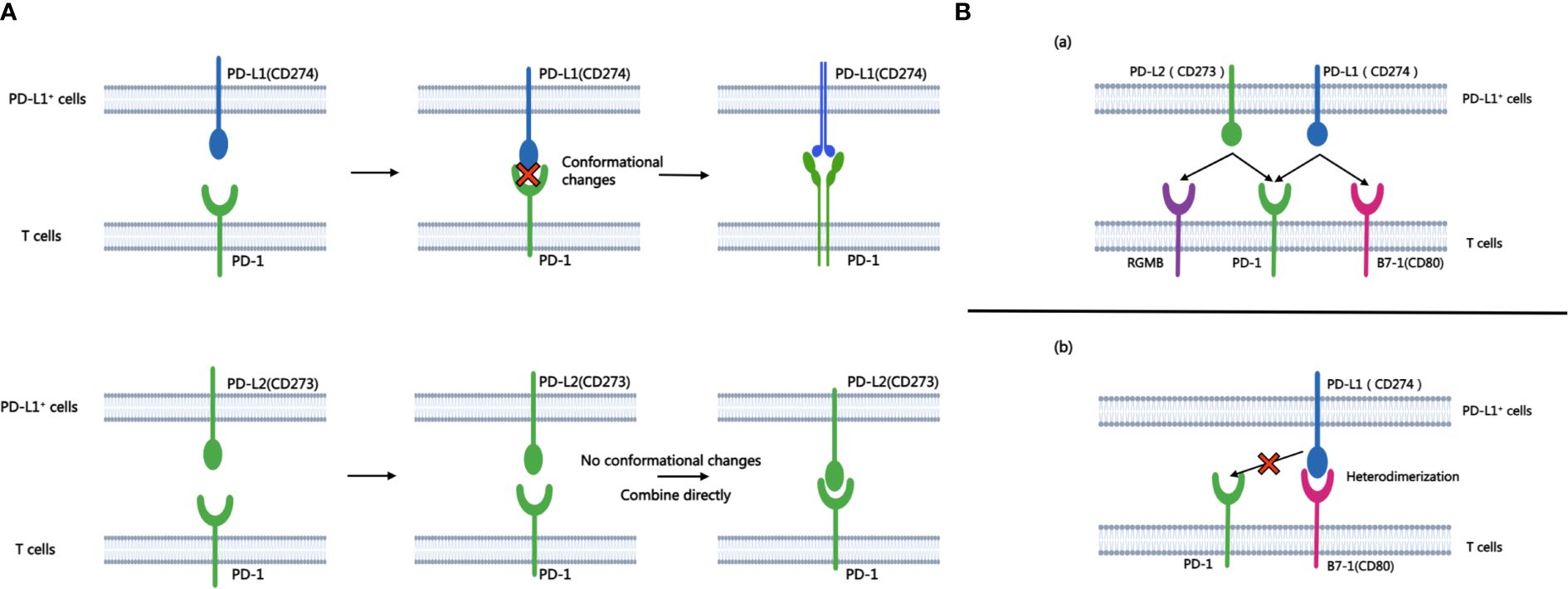
Figure 1 A shows the process of combining PD-L1 and PD-L2 with PD-1 respectively. B.(A) Besides PD-1, another binding site of PD-L1 and PD-L2 is PD-L2-RGMB and PD-L1-CD80. (B) CD80 can heterodimerize with PD-L1, which prevents the interaction between PD-L1 and PD-1.
This paper reviews the current status of PD-L2 research in tumors, encompassing the expression of PD-L2 in various common tumors, the association between PD-L2 expression and prognosis, regulatory factors, and the interplay between PD-L2 and tumor treatment.
2 Expression of PD-L2 in tumors and its relationship with prognosisStudies have identified the expression of PD-L2 in various common tumors, including but not limited to lung cancer, colorectal cancer, gastric cancer, esophageal cancer, head and neck cancer, breast cancer, cervical cancer, and liver cancer. High expression of PD-L2 tends to be associated with poor prognosis (Table 1).
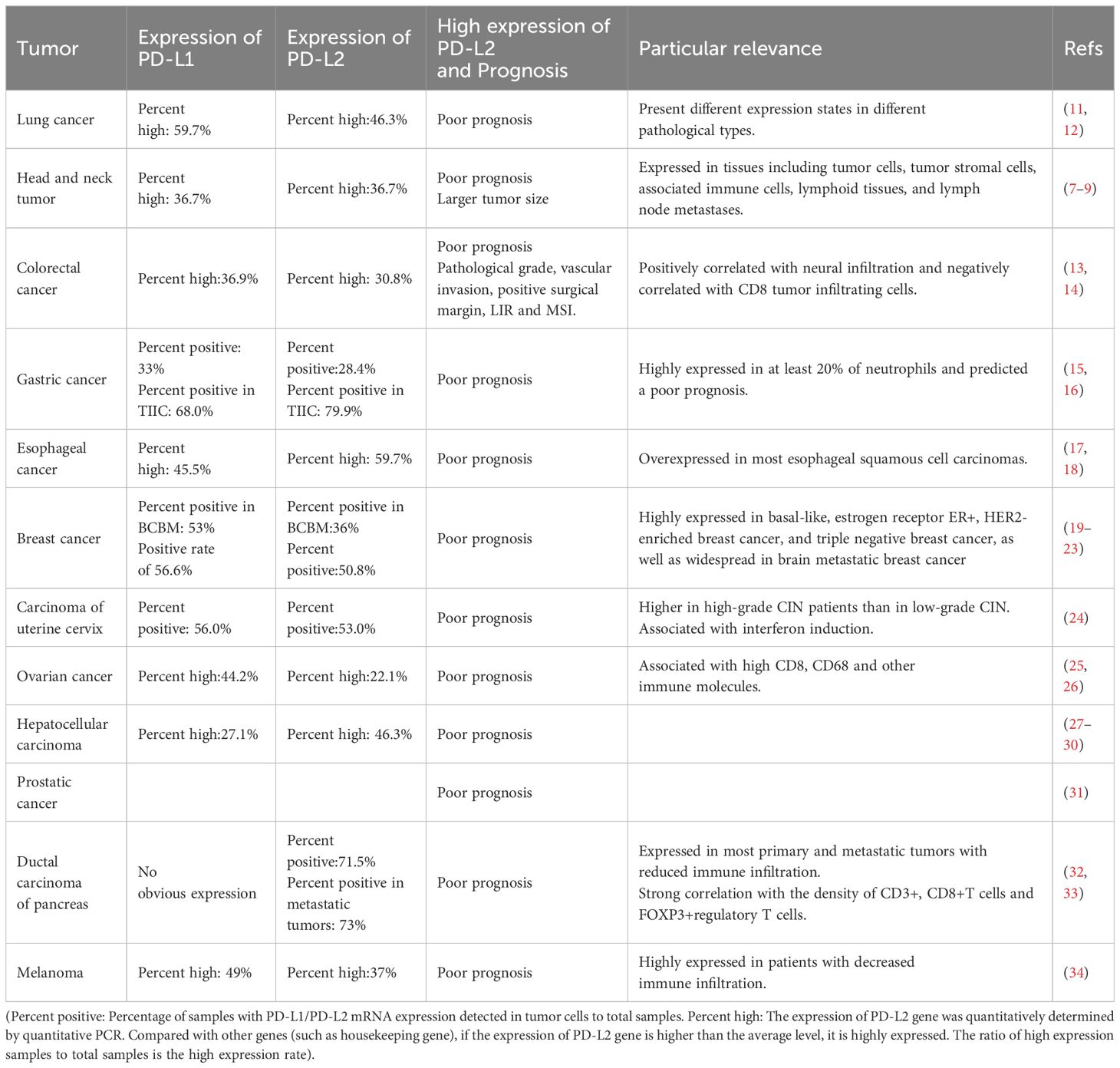
Table 1 The expression of PD-L2 in tumors and its association with clinical prognosis.
3 The regulatory network of the PD-L23.1 Signaling pathwaysIn tumor cells, the PD-L2 can be regulated by multiple signaling pathways, playing an important role in the occurrence and development of tumors. Therefore, it is imperative to devote sufficient attention to these signaling pathways to enhance our comprehension of the role of PD-L2 in tumorigenesis.
3.1.1 JAK/STAT signaling pathwayThe JAK/STAT pathway, formally recognized as the Janus kinase/signal transducer and transcriptional activator signaling pathway, stands as a pivotal communication hub in cellular function. An increasing body of evidence suggests that aberrations in the JAK/STAT pathway are linked to various cancers and autoimmune diseases (35). Studies have reported that the JAK/STAT pathway can trigger the expression of PD-L2 in tumor-related macrophages in lung adenocarcinoma. Additionally, soluble factors derived from cancer cells can induce the overexpression of PD-L2 in macrophages (36), hereby promoting tumor initiation and progression. Overexpression of PD-L1/PD-L2 in some malignant lymphomas is further linked to the activation or abnormalities of the JAK/STAT pathway, often accompanied by alterations in chromosome 9p24.1 (37, 38). These findings substantiate the role of the JAK/STAT pathway in regulating PD-L2 expression.
3.1.2 WNT signaling pathwayWNT (Wingless/Integrated) signaling serves as a crucial molecular regulator guiding a spectrum of physiological processes, encompassing embryonic development, adult stems cell homeostasis, and tissue regeneration (39, 40). Aberrations in the WNT signaling pathway are associated with the development and dysfunction of immune cells, as well as the promotion of immune escape (41). In tumor cells exhibiting high PD-L2 expression, a concomitant reduction in immune infiltration was often observed. In genetic identification, overexpression of genes within the WNT signaling pathway significantly correlated with the lack of immune infiltration (32). In melanoma, PD-L2 expression is also significantly increased in tumor cells with high phosphorylated β-catenin(pS1-βcat protein) expression downstream of the WNT/β-catenin signaling pathway (42), and the strong correlation between the high expression of this downstream protein and PD-L2 may offer insights for future tumor treatments targeting the signaling pathway.
3.1.3 NF-κB signaling pathwayThe transcription factor Nuclear factor-κB(NF-κB) plays a pivotal role in inflammation, oncogenesis, and tumor progression, being aberrantly activated in the majority of cancers, thereby contributing to tumorigenesis and progression (43). In a mouse liver cancer model, prolonged indomethacin use was observed to upregulate the expression of PD-L1 and PD-L2 through the TRIF/NF-κB and the JAK/STAT1 pathway. This effect ultimately led to a poor prognosis for hepatocellular carcinoma (HCC) by suppressing Tumor necrosis factor-α(TNF-α) and Interferon gamma-γ(IFN-γ) in liver cancer cells (44). Similarly, an investigation into the pathogenesis of systemic lupus erythematosus identified two crucial pathways, Toll-like receptor and type I interferon, which play a significant role in the development of Systemic lupus erythematosus. These pathways regulate the expression of PD-1 and its ligands (PD-L 2, PD-L1) through the activation of NF-κB, thereby promoting the occurrence and development of systemic lupus erythematosus (45). Furthermore, genetic analysis of clear cell renal carcinoma also revealed an association between the expression of PD-L2 and mRNA levels of NF-κB p65 (46).
3.1.4 AKT/mTOR signaling pathwayProtein kinase B/mammalian target of rapamycin (PI3K/Akt/mTOR) signaling pathway is a fundamental regulator of key physiological processes, including cell growth, migration, survival, and metabolism. Its dysregulation, a prevalent anomaly in tumor patients, significantly contributes to the initiation and progression of tumors (47). In a study focused on gene mutations in meningiomas, heightened PD-L2 expression was observed in patients with mutations in the AKT/mTOR signaling pathway when compared to other immune checkpoint proteins (48). In addition, Irina et al. underscored the pivotal role of PD-L2 in metastatic tumors, emphasizing its significance in tumor progression. They found that the level of the VHL protein, a tumor suppressor gene, determined the invasion capacity of tumors. By strategically targeting VHL through pathological means, they induced changes in the transcriptional and AKT/mTOR regulatory pathways, thereby influencing the expression of PD-L2. This intricate interplay ultimately fuels the initiation and progression of tumors (46, 49).
3.1.5 TLR9 signaling pathwayKnown as Toll-like receptor 9, is a key receptor for DNA recognition in cells, activating both innate and acquired immune responses and playing a crucial role in the immune system (50). Ongoing reports and clinical studies suggest promising potential for TLR9 agonists in cancer treatment (51). Baruah et al. reported that high expression of PD-L1/PD-L2 in fibroblasts from HPV-positive head and neck tumor patients is mediated by TLR9, which reduces the expression of PD-L1/PD-L2 when using the TLR9 specific antagonist, ODNTTAGGG (52). This finding opens a new path for targeted TLR9 to regulate immune checkpoint PD-1 and its ligands, which may be of important significance for the treatment of tumors. Moreover, this observation hints at a potential role for HPV in the intricate interplay of TLR9 and immune checkpoint modulation.
3.2 Genome and transcriptionAt the level of genome regulation, the amplification, deletion, or mutation of PD-L2-related genes will change the expression of PD-L2 at the source. The amplification of 9p24.1 will increase the transcription of the PD-L2 gene through Janus kinase 2(JAK2)/signal transduction and transcription activator 1(STAT1) signaling pathway (53). PD-L1-L2-SE, a super-enhancer located between the genes of CD274 and CD273, can induce and activate the transcription of PD-L2 (54). Methylation of the PD-L2 gene reduces the production of PD-L2 protein by inhibiting transcription. Histone acetylation will change the contact between histone and DNA and increase the transcription of the PD-L2 gene. Myc and HOXC10 are transcription-stimulating factors, and combining with the promoter region of PD-L2 gene will significantly increase the transcription level of PD-L2. Ganetespib, located upstream of Myc, suppresses the activity of heat shock protein 90 (HSP90), resulting in an alteration of PD-L2 expression (55). E26 transformation-specific variant transcription factor (ETV4) belongs to the ETS transcription factor family, which enhances PD-L2 transcription by binding to the PD-L1-L2-SE region. When octamer binding protein 2 (OCT2) binds to PD-L2 intron, OCT2 can increase the localization of PD-L2 on the B cell membrane. These regulatory sites may become potential targets for tumor immunotherapy (56).
3.3 Non-coding RNAThe majority of RNA transcribed from human genes cannot encode proteins, constituting what is known as non-coding RNAs. Despite their inability to code for proteins, these non-coding RNAs, including microRNA(miRNA), Long non-coding RNA(LncRNA), intron RNA, and repetitive RNA, exert significant influence on normal gene expression and contribute to various aspects of disease development. This study delves into the intricate impact of non-coding RNAs on the PD-L2, providing insights that highlight their potential as therapeutic targets for anti-tumor drugs (57). within this context, particular emphasis is placed on summarizing and elucidating the nuanced effects of miRNA and lncRNA on PD-L2 (Figure 2).
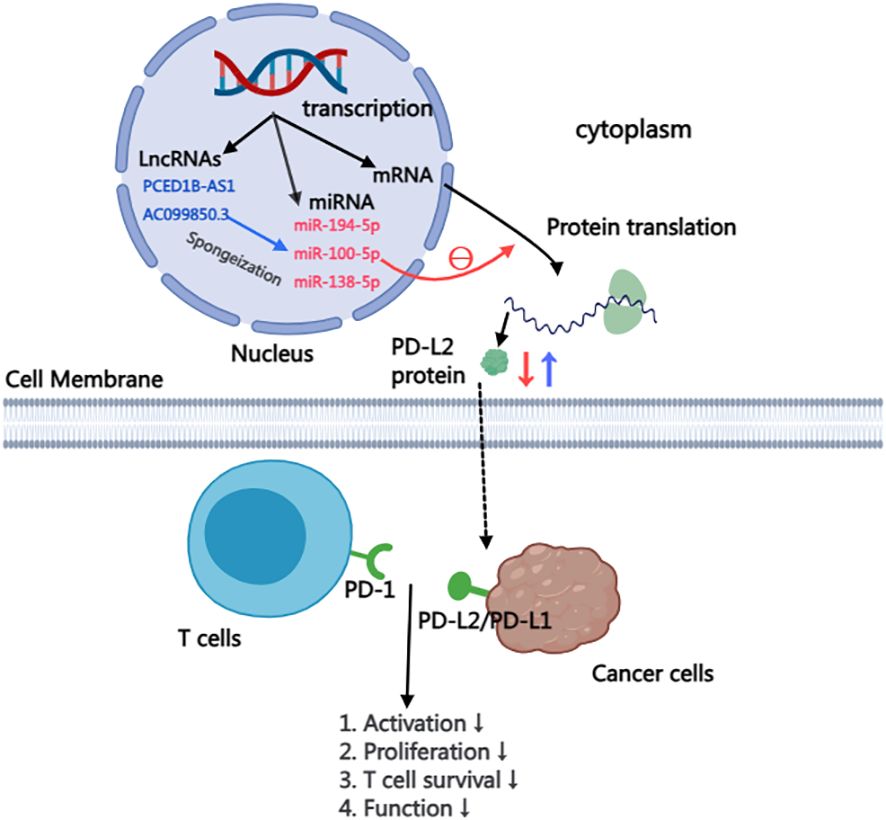
Figure 2 Mechanism of LncRNA and miRNA affecting PD-L2 expression in cells.
3.3.1 MiRNAMicroRNA, a subtype of non-coding RNA with a compact length ranging from 19 to 25 nucleotides, exerts its functional impact through post-transcriptional silencing of target genes. Functionally, miRNA plays a pivotal role in post-transcriptional silencing of target genes (58). This discussion succinctly summarizes the current understanding of miRNAs, focusing specifically on their roles in regulating the PD-L2, as detailed in Table 2.
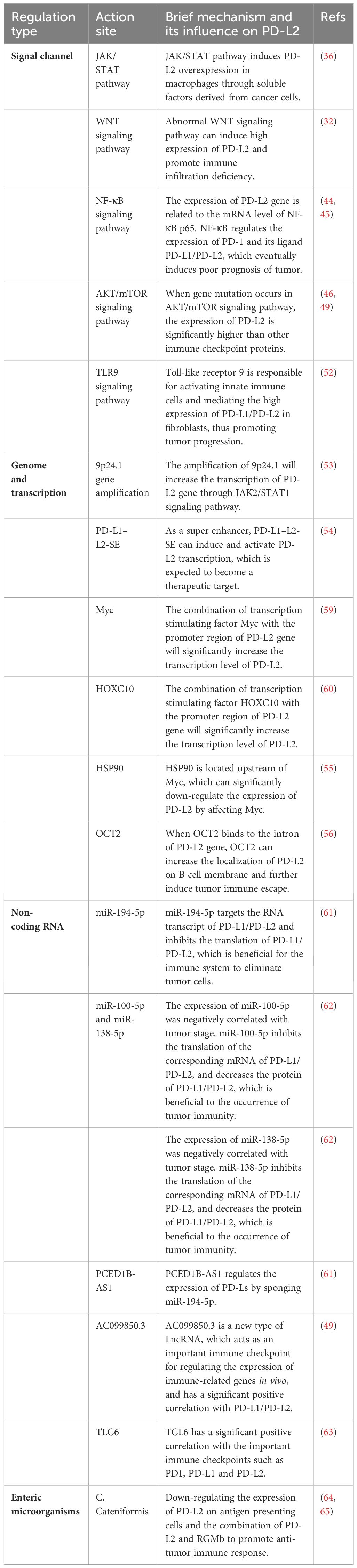
Table 2 Summary of factors regulating PD-L2.
MiR-194-5p: Plays a crucial role in the onset and advancement of various malignant tumors, encompassing HCC, renal cell carcinoma, melanoma, and pancreatic carcinoma (66–68). Mechanistically, miR-194-5p predominantly targets the RNA transcripts of PD-L1/PD-L2, hindering the translation of PD-L2 mRNA and consequently diminishing the protein content of PD-L2. Experimental evidence presented by Fan et al. empha sizes that hsa-mir-194-5p has the ability to interact simultaneously with the RNA transcripts of both PD-L1 and PD-L2. Through the use of miR-194-5p mi mics and inhibitors, it was observed that inhibiting miR-194-5p led to an increase in the protein levels of PD-L1/PD-L2, while miR-194-5p mimics resulted in a decrease in the protein levels of PD-L1/PD-L2. These findings distinctly indicate that miR-194-5p exerts targeted action on PD-Ls, inhibiting the translation of PD-1 ligand proteins. Consequently, in cases of abnormal function or absence of miR-194-5p, an overproduction of PD-L2 protein occurs. The interplay between PD-1 and PD-L2 induces immunosuppression in hepatocellular carcinoma, thereby fostering the initiation and progression of the tumor (61).
MiR-100-5p and miR-138-5p: Studies have shown that miR-100-5p and miR-138-5p are related to mitogen-activated protein kinase (MAPK) signaling pathway, apoptosis, and tumor necrosis factor (TNF) pathway, all of which are the regulators of cancer development, progression, and immune escape (62). In a study conducted by EI Ahanidi et al., the investigation into the relationship between miR-100-5p, miR-138-5p, and the TERT/PDL1/PD-L2 axis demonstrated that miR-138-5p exhibited elevated expression in normal bladder tissue, displaying a consistent downward trend from low-grade to high-grade tumors and from early-stage to progressive tumors. Notably, both miRNAs showed a negative correlation with tumor staging in bladder cell lines, and this negative correlation extended to the expression of PD-L1/PD-L2 protein (62). These results indicated that the combination of miR-100-5p and miR-138-5p to target genes inhibited their mRNA translation, resulting in the decrease of the expression level of PD-L1/PD-L2 protein, and finally induced immunosuppression of the tumor, further promoting the malignant progression and poor prognosis of the tumor.
3.3.2 LncRNALong non-coding RNA, defined by a length exceeding 200 nucleotides, plays a pivotal role in diverse biological processes. As a central focus in genetic research, LncRNA has been increasingly recognized as a critical regulatory factor for protein-encoding genes in various diseases, particularly cancer, where it significantly contributes to both the initiation and progression of tumor (69). Recent studies delving into the intricate network between lncRNA, and microRNA have revealed their regulatory role in immune checkpoint gene expression in breast cancer. This suggests that LncRNA may influence tumor progression by modulating immune checkpoint pathways (70).
PCED1B-AS1: Has been implicated in the pathogenesis of various human cancers, including gastric cancer, colorectal cancer, pancreatic ductal cancer, and renal clear cell carcinoma (71–74). Recent investigations highlight the role of lncRNA in regulating multiple target genes by acting as a miRNA sponge, effectively inhibiting miRNA function. Given the crucial role of PD-L1/PD-L2 in mediating tumor immunosuppression, Fan et al. reported that the long-chain non-coding RNA PCED1B-AS1 may function as a cytoplasmic sponge, modulating the expression of PD-Ls by sequestering miR-194-5p. Knocking down PCED1B-AS1 using lentivirus shRNA in a liver cancer cell line resulted in a simultaneous reduction in the protein levels of PD-L1 and PD-L2. Conversely, lentivirus overexpression of PCED1B-AS1 increased the protein levels of PD-L1 and PD-L2. These findings suggest that PCED1B-AS1 exerts control over the expression of PD-L1/PD-L2 in hepatoma cells by inhibiting miRNA function. Aberrations in PCED1B-AS1 may induce immunosuppression in hepatocellular carcinoma, ultimately contributing to tumor deterioration and progression (61).
AC099850.3: lncRNA-AC099850.3 emerges as a novel player with abnormal expression across various tumor types, including lung adenocarcinoma and hepatocellular carcinoma, showing associations with tumor staging, poor prognosis, and immune infiltration (75, 76). In HCC, the knockdown of AC099850.3 significantly impairs the in vitro proliferation and invasion capabilities of HCC and inhibits tumor cell growth in vivo. Notably, AC099850.3 exhibits a marked positive correlation with PD-L1 and PD-L2, key immune checkpoints in vivo. High expression of PD-L1/PD-L2 is generally linked to poor tumor prognosis, indicating that AC099850.3 functions as an immune-related gene, regulating the expression of critical immune checkpoints in HCC. This, in turn, induces immunosuppression in HCC. Furthermore, studies have revealed that AC099850.3 promotes the malignant progression of hepatocellular carcinoma through the AKT signaling pathway. Spirina et al. reported that the AKT signaling pathway exerts a regulatory effect on PD-L2 (49).
TLC6: T-cell leukemia/lymphoma 6, a novel regulator implicated in HCC and renal clear cell carcinoma progression, associated with a poor prognosis (77, 78). In breast cancer, low TCL6 expression independently predicts an adverse outcome for PR (estrogen and progesterone receptor)-negative patients, while also correlating with immune infiltrating cells, including B cells, CD4+ T cells, and CD8+ T cells. Furthermore, TCL6 shows a significant positive correlation with PD1, PD-L1, and PD-L2 (63). However, the pathway or mechanism by which TCL6 acts on the expression of PD-L1/PD-L2 still needs further study (20). TCL6 presents a promising avenue for exploration in cancer therapeutics.
3.4 Gut microbiomeA large number of studies have shown that microorganisms in the intestinal tract and other parts may promote the occurrence and development of cancer, and affect cancer immune surveillance and response to immunotherapy (79). Studies have reported that intestinal microflora can down-regulate the expression of PD-L2 in CD11c+DCs (dendritic cells) in the intestine and tumor drainage lymph nodes and can promote anti-tumor immunity by combining with partner repulsion guiding molecule B (RGMB). Park et al. discovered an intestinal flora-C. cateniformis, which can induce the above-mentioned effects. The combination of PD-L2 and RGMB plays a role in inhibiting anti-cancer T cells. Blocking the interaction between PD-L2 and RGMB with antibodies can further relieve the inhibition of T cells, which is beneficial for T cells to clear cancer cells. This indicates that PD-L2 not only inhibits T cells through PD-1 but also exerts inhibitory effects through RGMB (64, 65). This also confirmed a new strategy of cancer immunotherapy-blocking the interaction between PD-L2 and RGMB to enhance the response to cancer immunotherapy.
4 Treatment strategy4.1 Small molecular drugsBecause the affinity of PD-1 to PD-L1 and PD-1 to PD-L2 is quite different. These small molecules hold the potential to penetrate the tumor microenvironment more effectively than traditional anti-PD-1/PD-L1 axis drugs, such as pabolizumab, potentially enhancing therapeutic efficacy (80). One notable molecular targeted drug in this context is JQ1, a BET-bromine domain inhibitor. JQ1 inhibits the binding of the BET-bromine domain and histone, suppressing the transcription of target genes. In studies conducted by Liu et al., JQ1 treatment led to a reduction in PD-L2 mRNA levels in renal cell carcinoma, and prostate, liver, and lung cancer cell lines (81). Therefore, JQ1 may be a potential molecular drug for tumor treatment. Table 3 summarizes the current new tumor treatment strategies for PD-L2. Although some of them are still in the research stage, they represent the new direction of tumor treatment in the future.
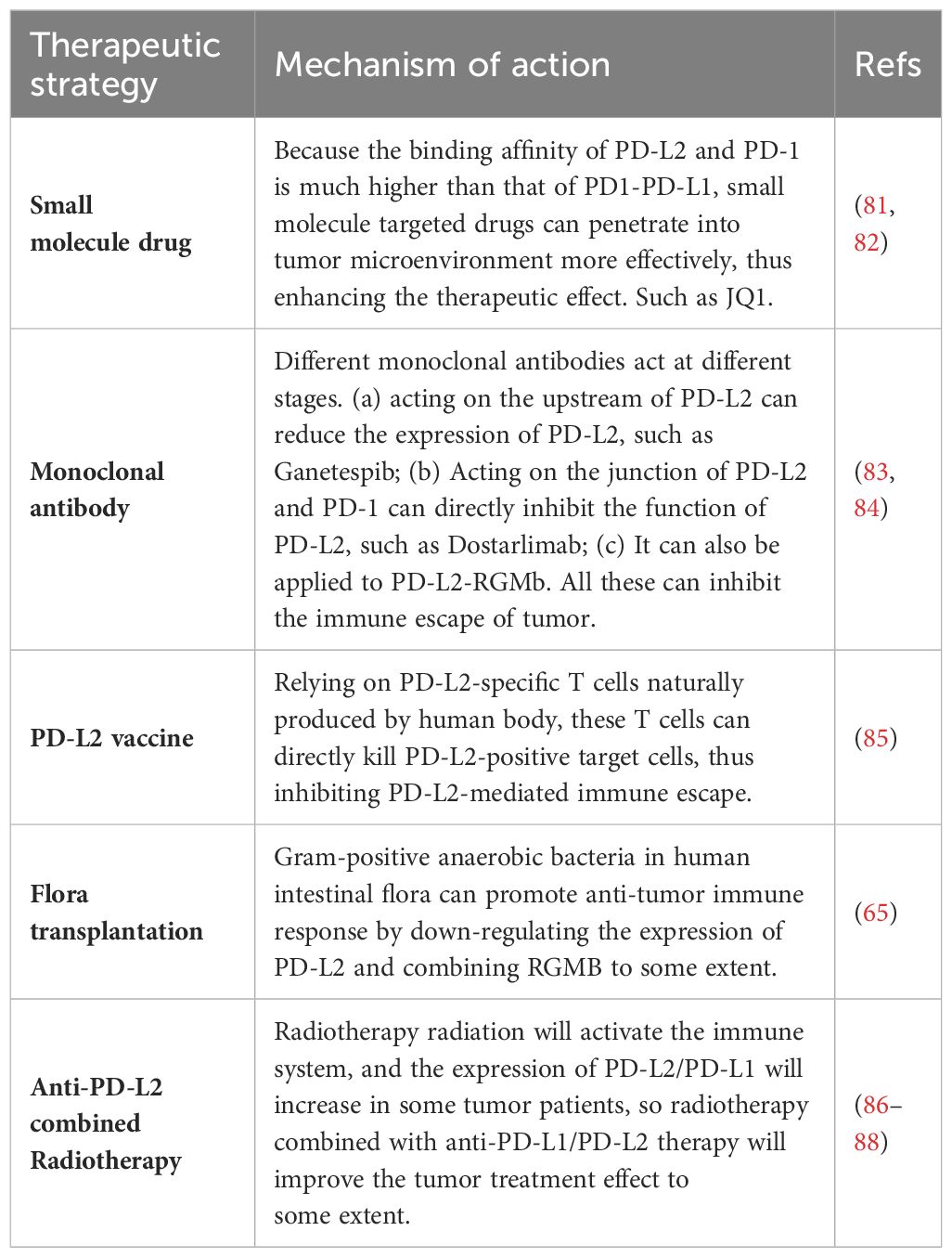
Table 3 Tumor treatment strategy for PD-L2.
4.2 PD-L2 vaccineThe so-called PD-L2 vaccine is to introduce PD-L2 antigen or proteins or cells carrying the restricted epitopes of human leukocyte antigen (HLA) expressed by PD-L2 into patients, thus inducing the activation of PD-L2 specific T cells. PD-L2-specific T cells, naturally produced in cancer patients, play a dual role in supporting anti-tumor immunity. These T cells can directly kill target cells and indirectly contribute to anti-tumor immunity by releasing pro-inflammatory factors in the tumor microenvironment. In clinical trials, researchers aim to enhance the immune response against PD-L2 by administering the PD-L2 vaccine, hoping to complement other forms of immunotherapy. While the treatment’s effectiveness hasn’t shown significant improvement in patients, it holds potential as a target for cancer vaccine and immune checkpoint-blocking treatments (85).
4.3 Combination with radiotherapyAs a vital component of tumor treatment, the potential of radiotherapy interactions with immune checkpoint inhibitors are also reported. A study revealed a significant increase in PD-L1 and PD-L2 expression in MCF-120 (human breast cancer cells) after hyperthermia (HT) and radiotherapy RT (2×5 Gy) treatment at any temperature (86). Considering the elevated expression of immune checkpoint molecules, incorporating immune checkpoint inhibitors into the combined tumor hyperthermia and radiotherapy approach may optimize therapeutic outcomes. In addition, radiotherapy is previously reported to be able to locally activate the immune system, potentially inducing a systemic immune response and targeting distant metastatic tumors (87). Previous research demonstrated increased efficacy of paolizumab in lung cancer patients who underwent prioradiotherapy (88). And the activation of immune system by radiotherapy radiation may be optimized and improved in tumors with more inhibition of immune system by PD-L2 at baseline. However, understanding the pathway through which radiotherapy affects PD-L1/PD-L2 and investigating the potential relationship between the immune response induced by local radiotherapy require further exploration (31).
4.4 Intestinal flora transplantationPrevious studies have shown that transplantation of fecal flora from patients with good response to PD-1 therapy can reduce the drug resistance of melanoma patients to PD-1 therapy, thus improving the tumor treatment effect. Gram-positive anaerobic bacteria in human intestinal microorganisms can promote anti-tumor immune response by down-regulating the expression of PD-L2 and binding RGMB to some extent (65). This will further expand the starting point of tumor treatment. On the one hand, it inspires us to fully strengthen the study of intestinal flora, clarify the types of intestinal microorganisms for tumor treatment, and bring a new dawn to tumor patients who have poor responses to immunotherapy through targeted flora transplantation. On the other hand, using intestinal microorganisms as a discovery platform to identify a new target of cancer immunotherapy-PD-L2-RGMB, and blocking the interaction between PD-L2 and RGMB can enhance the response to cancer immunotherapy. Small molecule drugs targeting specific intestinal microorganisms can be further developed to enhance the effect of cancer immunotherapy.
5 ProspectsIn conclusion, PD-L2 stands as a key player in various human cancers, showing significant promise as a therapeutic target. Currently, there exist numerous constraints, notably the incomplete comprehension of PD-L2 expression and its regulatory mechanism, as well as the insufficient exploration of related signal pathways. Moreover, the clinical approval of anti-PD-L2 antibodies for tumor treatment remains elusive, necessitating further investigation and discovery. However, the high affinity of PD-L2 for PD-1, the new insight that intestinal microorganisms regulate PD-1 pathway, and the richness of PD-L2 related therapies all indicate that cancer immunotherapy is expected to make a breakthrough. The discovery of new pathways such as PD-L2-RGMB has further expanded the scope of targeted therapy. The continuous exploration of clinical application of PD-L2 has brought exciting prospects for promoting cancer treatment. The full names of abbreviations in this article are displayed in Table 4.
Table 4 Summary of abbreviations.
Author contributionsYY: Writing – original draft, Writing – review & editing. XY: Writing – original draft, Writing – review & editing. XB: Conceptualization, Writing – review & editing. JY: Investigation, Writing – review & editing. JS: Writing – original draft, Writing – review & editing.
FundingThe author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References1. Wang J, Seebacher N, Shi H, Kan Q, Duan Z. Novel strategies to prevent the development of multidrug resistance (MDR) in cancer. Oncotarget. (2017) 8:84559–71. doi: 10.18632/oncotarget.v8i48
PubMed Abstract | CrossRef Full Text | Google Scholar
3. Salmaninejad A, Khoramshahi V, Azani A, Soltaninejad E, Aslani S, Zamani MR, et al. PD-1 and cancer: molecular mechanisms and polymorphisms. Immunogenetics. (2018) 70:73–86. doi: 10.1007/s00251-017-1015-5
PubMed Abstract | CrossRef Full Text | Google Scholar
4. Yearley JH, Gibson C, Yu N, Moon C, Murphy E, Juco J, et al. PD-L2 expression in human tumors: relevance to anti-PD-1 therapy in cancer. Clin Cancer Res. (2017) 23:3158–67. doi: 10.1158/1078-0432.CCR-16-1761
PubMed Abstract | CrossRef Full Text | Google Scholar
5. Ghiotto M, Gauthier L, Serriari N, Pastor S, Truneh A, Nunès J, et al. PD-L1 and PD-L2 differ in their molecular mechanisms of interaction with PD-1. Int Immunol. (2010) 22:651–60. doi: 10.1093/intimm/dxq049
PubMed Abstract | CrossRef Full Text | Google Scholar
6. Youngnak P, Kozono Y, Kozono H, Iwai H, Otsuki N, Jin H, et al. Differential binding properties of B7-H1 and B7-DC to programmed death-1. Biochem Biophys Res Commun. (2003) 307:672–7. doi: 10.1016/S0006-291X(03)01257-9
PubMed Abstract | CrossRef Full Text | Google Scholar
7. Lázár-Molnár E, Scandiuzzi L, Basu I, Quinn T, Sylvestre E, Palmieri E, et al. Structure-guided development of a high-affinity human Programmed Cell Death-1: Implications for tumor immunotherapy. EBioMedicine. (2017) 17:30–44. doi: 10.1016/j.ebiom.2017.02.004
PubMed Abstract | CrossRef Full Text | Google Scholar
8. Butte MJ, Keir ME, Phamduy TB, Sharpe AH, Freeman GJ. Programmed death-1 ligand 1 interacts specifically with the B7-1 costimulatory molecule to inhibit T cell responses. Immunity. (2007) 27:111–22. doi: 10.1016/j.immuni.2007.05.016
PubMed Abstract | CrossRef Full Text | Google Scholar
9. Xiao Y, Yu S, Zhu B, Bedoret D, Bu X, Francisco LM, et al. RGMb is a novel binding partner for PD-L2 and its engagement with PD-L2 promotes respiratory tolerance. J Exp Med. (2014) 211:943–59. doi: 10.1084/jem.20130790
PubMed Abstract | CrossRef Full Text | Google Scholar
10. Burke KP, Chaudhri A, Freeman GJ, Sharpe AH. The B7:CD28 family and friends: Unraveling coinhibitory interactions. Immunity. (2024) 57:223–44. doi: 10.1016/j.immuni.2024.01.013
PubMed Abstract | CrossRef Full Text | Google Scholar
11. Ludovini V, Bianconi F, Siggillino A, Vannucci J, Baglivo S, Berti V, et al. High PD-L1/IDO-2 and PD-L2/IDO-1 co-expression levels are associated with worse overall survival in resected non-small cell lung cancer patients. Genes. (2021) 12(2):273. doi: 10.3390/genes12020273
PubMed Abstract | CrossRef Full Text | Google Scholar
12. Takamori S, Takada K, Azuma K, Jogo T, Shimokawa M, Toyokawa G, et al. Prognostic impact of programmed death-ligand 2 expression in primary lung adenocarcinoma patients. Ann Surg Oncol. (2019) 26:1916–24. doi: 10.1245/s10434-019-07231-z
PubMed Abstract | CrossRef Full Text | Google Scholar
13. Huang KCY, Chiang SF, Chen TW, Chen WTL, Yang PC, Ke TW, et al. Prognostic relevance of programmed cell death 1 ligand 2 (PDCD1LG2/PD-L2) in patients with advanced stage colon carcinoma treated with chemotherapy. Sci Rep. (2020) 10:22330. doi: 10.1038/s41598-020-79419-3
PubMed Abstract | CrossRef Full Text | Google Scholar
14. Zengin M, Zergeroğlu S, Okcu O, Benek S. PD-1 and PD-L2 expression predict relapse risk and poor survival in patients with stage III colorectal cancer. Cell Oncol. (2021) 44:423–32. doi: 10.1007/s13402-020-00579-5
CrossRef Full Text | Google Scholar
15. Shan ZG, Zhao YL, Zhang JY, Yan ZB, Wang TT, Mao FY, et al. FasL+ PD-L2+ Identifies a novel immunosuppressive neutrophil population in human gastric cancer that promotes disease progression. Adv Sci Weinh Baden-Wurtt Ger. (2022) 9:e2103543. doi: 10.1002/advs.202103543
CrossRef Full Text | Google Scholar
16. Nakayama Y, Mimura K, Kua LF, Okayama H, Min AKT, Saito K, et al. Immune suppression caused by PD-L2 expression on tumor cells in gastric cancer. Gastric Cancer. (2020) 23:961–73. doi: 10.1007/s10120-020-01079-z
PubMed Abstract | CrossRef Full Text | Google Scholar
17. Tanaka K, Miyata H, Sugimura K, Kanemura T, Hamada-Uematsu M, Mizote Y, et al. Negative influence of programmed death-1-ligands on the survival of esophageal cancer patients treated with chemotherapy. Cancer Sci. (2016) 107:726–33. doi: 10.1111/cas.12938
PubMed Abstract | CrossRef Full Text | Google Scholar
18. Zhao JJ, Zhou ZQ, Wang P, Chen CL, Liu Y, Pan QZ, et al. Orchestration of immune checkpoints in tumor immune contexture and their prognostic significance in esophageal squamous cell carcinoma. Cancer Manag Res. (2018) 10:6457–68. doi: 10.2147/CMAR
PubMed Abstract | CrossRef Full Text | Google Scholar
19. Duchnowska R, Pęksa R, Radecka B, Mandat T, Trojanowski T, Jarosz B, et al. Immune response in breast cancer brain metastases and their microenvironment: the role of the PD-1/PD-L axis. Breast Cancer Res BCR. (2016) 18:43. doi: 10.1186/s13058-016-0702-8
CrossRef Full Text | Google Scholar
20. Baptista MZ, Sarian LO, Derchain SFM, Pinto GA, Vassallo J. Prognostic significance of PD-L1 and PD-L2 in breast cancer. Hum Pathol. (2016) 47:78–84. doi: 10.1016/j.humpath.2015.09.006
PubMed Abstract | CrossRef Full Text | Google Scholar
21. Chervoneva I, Peck AR, Sun Y, Yi M, Udhane SS, Langenheim JF, et al. High PD-L2 predicts early recurrence of ER-positive breast cancer. JCO Precis Oncol. (2023) 7:e2100498. doi: 10.1200/PO.21.00498
PubMed Abstract | CrossRef Full Text | Google Scholar
22. Solinas C, Garaud S, De Silva P, Boisson A, Van den Eynden G, de Wind A, et al. Immune checkpoint molecules on tumor-infiltrating lymphocytes and their association with tertiary lymphoid structures in human breast cancer. Front Immunol. (2017) 8:1412. doi: 10.3389/fimmu.2017.01412
PubMed Abstract | CrossRef Full Text | Google Scholar
23. Montoyo-Pujol YG, García-Escolano M, Ponce JJ, Delgado-García S, Martín TA, Ballester H, et al. Variable intrinsic expression of immunoregulatory biomarkers in breast cancer cell lines, mammospheres, and co-cultures. Int J Mol Sci. (2023) 24:4478. doi: 10.3390/ijms24054478
PubMed Abstract | CrossRef Full Text | Google Scholar
24. Rotman J, den Otter LAS, Bleeker MCG, Samuels SS, Heeren AM, Roemer MGM, et al. PD-L1 and PD-L2 expression in cervical cancer: regulation and biomarker potential. Front Immunol. (2020) 11:596825. doi: 10.3389/fimmu.2020.596825
PubMed Abstract | CrossRef Full Text | Google Scholar
26. Batman S, Matsuo K, Mhawech-Fauceglia P, Munro E, Weisenberger M, Allen A, et al. Intersection of DNA repair pathways and the immune landscape identifies PD-L2 as a prognostic marker in epithelial ovarian cancer. Cancers. (2021) 13:1972. doi: 10.3390/cancers13081972
PubMed Abstract | CrossRef Full Text | Google Scholar
27. Jung HI, Jeong D, Ji S, Ahn TS, Bae SH, Chin S, et al. Overexpression of PD-L1 and PD-L2 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Res Treat Off J Korean Cancer Assoc. (2017) 49:246–54. doi: 10.4143/crt.2016.066
CrossRef Full Text | Google Scholar
28. Yang H, Zhou X, Sun L, Mao Y. Correlation between PD-L2 expression and clinical outcome in solid cancer patients: A meta-analysis. Front Oncol. (2019) 9:47. doi: 10.3389/fonc.2019.00047
PubMed Abstract | CrossRef Full Text | Google Scholar
29. Shrestha R, Bridle KR, Crawford DHG, Jayachandran A. TNF−α−mediated epithelial−to−mesenchymal transition regulates expression of immune checkpoint molecules in hepatocellular carcinoma. Mol Med Rep. (2020) 21:1849–60. doi: 10.3892/mmr
PubMed Abstract | CrossRef Full Text | Google Scholar
30. Tao YY, Shi Y, Gong XQ, Li L, Li ZM, Yang L, et al. Radiomic analysis based on magnetic resonance imaging for predicting PD-L2 expression in hepatocellular carcinoma. Cancers. (2023) 15:365. doi: 10.3390/cancers15020365
留言 (0)