The extracellular matrix (ECM) is an intricate network of multiple macromolecules, consisting mainly of collagen, non-collagenous glycoproteins, amino glycan, proteoglycans, and elastin [1]. The ECM provides a microenvironment for cell survival and activity. It not only plays static physical roles in supporting, connecting, retaining water, and protecting cells, but also dynamically exerts a full range of biological effects on cells [2,3]. The ECM components regulate the activation of cell surface receptors. This affects the activation or inhibition of intracellular signaling pathways, which influences the expression and function of ncRNA [4,5]. The expression of ncRNA also affects changes in ECM components, thus participating in the molecular mechanisms that regulate ECM remodeling [6,7]. When the synthesis and metabolism of the ECM are imbalanced, the crosslinks between the ECM components increase and strengthen. Collagen and other proteins are subsequently deposited in the ECM, leading to changes in ECM stiffness and closer binding of integrins to their ligands. This activates a series of biological changes that have profound physiological and pathological implications [8,9].
Several studies have shown that ECM stiffness mechanically modulates cell behavior under physiological conditions. Engler et al. demonstrated that the physical properties of the local microenvironment play a key role in controlling cell differentiation [10]. In photopolymerizable methacrylamide chitosan (MAC) biomaterials, it was demonstrated that ECM stiffness exerts an important influence on the proliferation and differentiation of adult neural stem/progenitor cells (NSPC) [11]. The mechanism of action for the regulation of matrix stiffness during physiological development is complex and involves a variety of intracellular protein components. MiRNAs and lncRNAs play an important role in ECM modification [12].Of course, matrix stiffness also regulates cell migration and adhesion through cellular components such as integrins and actin [13,14]. Matrix stiffness affects cell-to-cell communication by regulating the fibronectin-adhesion patch kinase FAK-Cx43 (connexin 43) axis, which alters gap junction formation in a group of cells [15]. A recent study has revealed that in addition to actin, microtubules that make up the cytoskeleton also play a key role in altering nuclear morphology [16]. Mechanisms of glycolysis in response to the mechanical regulation of cytoskeletal structures were realized to correlate cellular metabolism with the mechanical properties of surrounding tissues [17,18]. Overall, ECM stiffness manipulated cell morphology [19], growth, differentiation [20], and metabolism [17], and regulated cell migration and adhesion [13,14]through cell shape and cytoskeletal remodeling during physiological development [20].
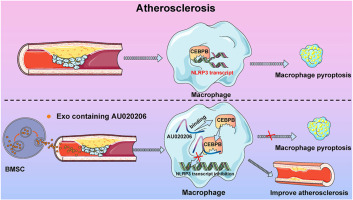
Indeed, previous studies have highlighted the crucial role of ECM stiffness in various pathological processes. For example, atherosclerosis, a disease characterized by the formation and progression of arterial plaques, is strongly associated with ECM stiffness. Remodeling of the ECM is a key factor in altering arterial stiffness, and many proteins and cytokines are involved in this process [21]. Matrix stiffness is also strongly associated with fibrotic diseases; matrix stiffness promotes cardiac fibroblast differentiation by enhancing POU2F1 expression through down-regulation of fibrosis inhibitory factors [22]. Mechanical stretching leads to the activation of TGF-β1 and the progression of fibrosis in fibrotic lung tissues [23], and matrix stiffness also regulates hepatic fibrosis [24] and disease progression in renal fibrosis [25]. Chondrocyte YAP activity is associated with ECM stiffening and osteoarthritis pathogenesis [26], and matrix stiffness affects keratinization [27]. ECM stiffness has also been shown to be important during cancer development. It was found that increased ECM hardness promotes hepatocellular carcinoma (HCC) cell proliferation [28]. Therefore, as an important physical property of the ECM, the hardness of the ECM plays a crucial role in regulating the growth, proliferation, motility, metabolism, and invasion of cancer cells [29].
Since ECM stiffness was associated with both physiological and pathological processes, the regulation mechanism was intriguing. Previous studies of ECM stiffness have disclosed several mechanosensing molecules. For example, Piezo1, a component of piezo-type mechanosensitive ion channels, has been found to be involved in the regulation of Drosophila glioblastoma stem cells. It was found to be overexpressed and associated with a malignant prognosis in gliomas. Piezo1 interacts with focal adhesion kinase (FAK) signaling to increase tissue stiffness, leading to promoted glioma cell proliferation and invasion [30]. Integrin β1 functions as a mechanical force signaling molecule that transmits changes in ECM stiffness to HCC cells and activates downstream molecular events [31].YAP and tafazzin (TAZ) were well-known mechanotransduction molecules. ECM stiffness regulated cellular behaviors by activating YAP/TAZ through both hippos-dependent and independent pathways [32]. Moreover, increased ECM stiffness also activated epithelial-mesenchymal transition (EMT) signaling through Snail, which affected the abscission mechanism of mammary epithelial cells [33].
Recently, it has been reported that non-coding RNA (ncRNA), which is not translated into proteins, makes important contributions to various biological processes and plays a crucial role in epigenetic regulation [34]. MicroRNAs (miRNAs) and long non-coding RNAs (lncRNAs) are two common types of ncRNAs. However, most well-known ECM stiffness-related RNAs belonged to messenger RNAs (mRNAs). Emerging evidence has revealed that ncRNAs like miRNAs and lncRNAs were frequently regulated by ECM stiffness in various diseases, including carcinogenesis and vascular diseases [35]. In this review, we have summarized the latest findings on miRNAs and lncRNAs associated with ECM stiffness and described how these ncRNAs affect disease processes. Furthermore, we have discussed the interactions between miRNAs and lncRNAs with ECM stiffness and integrated a signaling map in which these ncRNAs participate. Ultimately, we explored the perspectives and significance of investigations into ECM stiffness associated with ncRNAs.








留言 (0)