
記住我
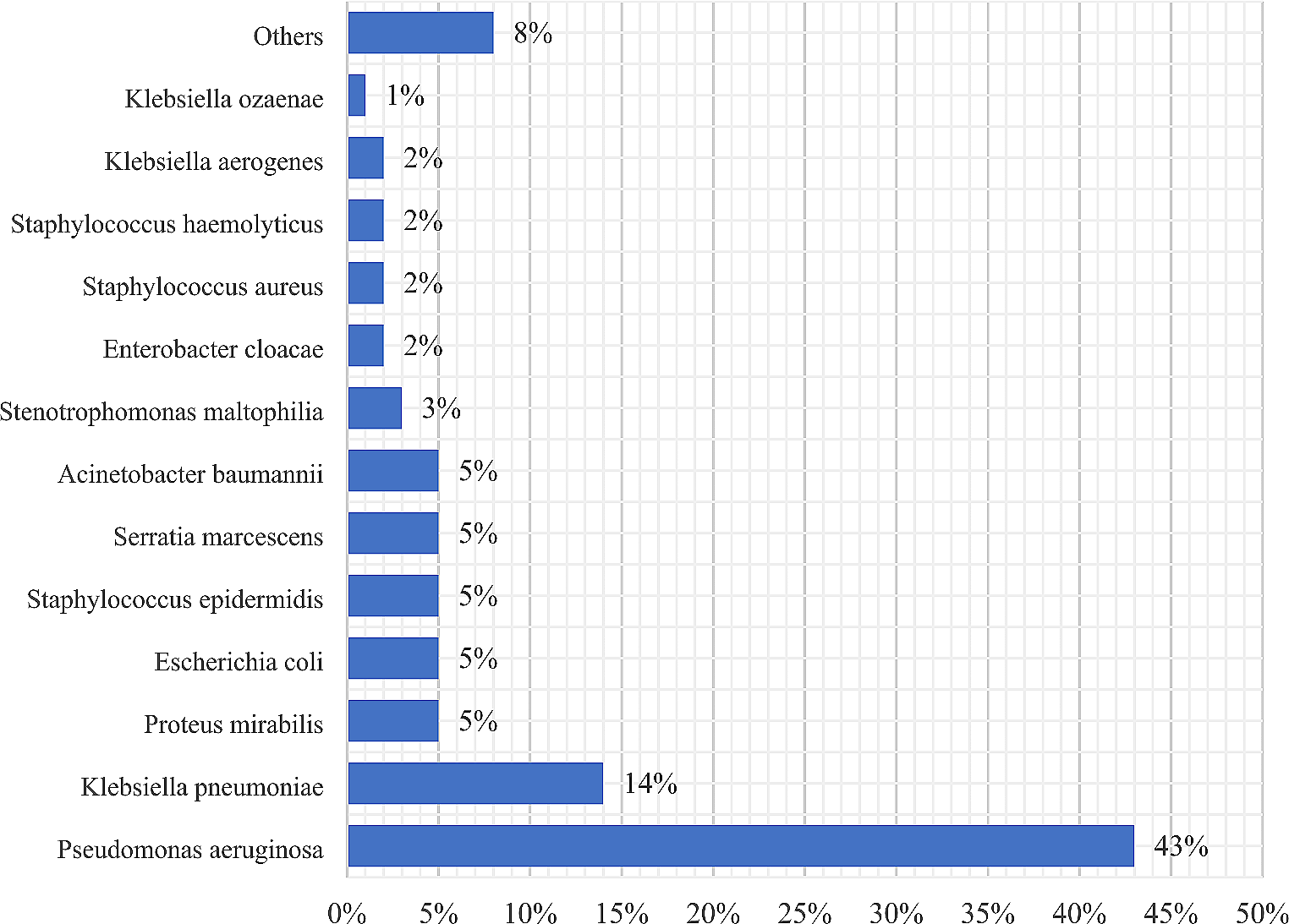

In this study, a collection of 236 clinical enterococci was analyzed for VVE detection. Eleven E. faecium strains (4.6% of all enterococcal isolates) were considered putative VVE (VVEIta-S) being vanA positive and vancomycin susceptible. In our hospital, the frequency of putative VVE (19%) among the E. faecium clinical isolates (n = 59) was higher than those reported in other studies and surveys [9, 16, 18], although a progressive increase of the VVE prevalence has been recently reported in several countries [6, 15, 16]. The 11 putative VVEIta-S were also tested for their susceptibility to ampicillin, ciprofloxacin, erythromycin, linezolid, tedizolid, and chloramphenicol, as summarized in Table 1. All the isolates were resistant to ampicillin and ciprofloxacin (MIC, > 128 mg/L) and ten to erythromycin (MIC, > 128 mg/L). All isolates were susceptible, besides to vancomycin, to linezolid (MIC, range 1–4 mg/L), tedizolid (MIC, range 0.5–1 mg/L), and chloramphenicol (MIC, range 8–16 mg/L).
Table 1 Distinctive features, susceptibility patterns, and molecular typing of the 11 vancomycin susceptible isolates were investigatedMolecular analysis of VVEIta-S strainsThe 11 putative VVEIta-S were typed by PFGE and MLST to establish their genetic relatedness (Table 1).
PFGE clustering showed two main clones: the first one included the 700,907, 727,475, 749,325, and 749,286 isolates exhibiting the same PFGE pattern (pulsotype A) and the closely related 741,160 isolate (A1 pulsotype). The second clone was represented by the 732,558 isolate showing the pulsotype B and the closely related 733,387 and 731,980 strains with pulsotypes B1 and B2, respectively. The 755,686, 742,783, and 735,902 isolates belonged to three different pulsotypes: C, D, and E, respectively (Figure S1).
By MLST five strains, associated to pulsotype A and A1 (700,907, 749,325, 749,286, 727,475, and 741,160), belonged to the newly reported pstS-null sequence type 1478 (ST1478). To the best of our knowledge, this is the first report of VVE-S included in this particular clonal lineage. Previously, VVE strains lacking the pstS gene have been identified only in the ST1421 clone [6, 10, 16]. To date, VRE ST1478 was only detected across Canadian hospitals by national surveillance [24, 25].
The isolates included in the B, B1, B2, and C pulsotypes belonged to ST80, whereas the 742,783 (pulsotype D) and 735,902 (pulsotype E) isolates were associated to ST117 and ST789, respectively (Table 1). Many of the more recently dominant E. faecium sequence types associated with nosocomial infection, such as ST80, ST789, and ST117, are derived from the well-known clonal complex CC17 [33]. Interestingly, the ST789 has been reported for clinical vancomycin-resistant E. faecium isolates in Algeria [34]. To date, most of the VVE described in the literature are included in the ST1421, ST203, and ST18 [6, 9,10,11, 14, 16], different from those found in this study (ST117 and ST80 (Table 1)), probably due to VVEIta-S evolution from the most common VRE circulating clones in our hospital [26].
Genome analysis of VVEIta-S strainsOn the basis of PFGE results, one strain for each pulsotype was subjected to WGS, resulting in a total of eight isolates sequenced (Table 2).
Table 2 Main features of the 7 VVEIta-S strains and the E. faecium 741,160 subjected to WGSResFinder analysis of the eight genomes revealed complex resistomes for the presence of several acquired antibiotic resistance genes (Table 2). All isolates shared, besides vanH/vanA/vanX cluster, msrC (resistance to macrolides and streptogramins A) and aac(6′)-Ii (resistance to aminoglycosides). The erm(B) (resistance to macrolides, lincosamides, and streptogramins A) and ant(6)-Ia, aph(3′)-III, and aac(6′)-aph(2″) (resistance to aminoglycosides) were identified in all but two isolates (732,558 and 735,902). Three out of eight genomes exhibited the dfrG (resistance to trimethoprim) and tet(M) (resistance to tetracycline) genes, and only the 755,686 genome showed the double combination tet(M)/tet(L).
The study of the virulome revealed that the eight isolates shared three acquired virulence genes: acm (collagen-binding protein), efaAfm (gelatinase), and hylEfm (hyaluronidase). The espfm (enterococcal surface protein biofilm associated to biofilm production) was identified in all but two isolates (742,783 and 735,902).
In silico analysis of WGS data showed that the 8 isolates exhibited different SNPs (from a minimum of 18 to a maximum of 3762), as detailed in Table S1. The clonal relatedness among strains is also shown in Figure S2.
An in-depth analysis of the 700,907 and 741,160 genomes, belonging to the pstS-null ST1478, clarified the absence of the housekeeping gene. Indeed, this gene was truncated by the insertion of a 2894 bp region, containing two ISL3 family transposases, causing the loss of 503 bp of the pstS gene. However, despite the missing pstS in these two genomes, sequencing analysis revealed a pstS homologue within a pst operon (also referred to as pstS2), which is thought to be the actual pstS housekeeping gene in E. faecium [22].
Moreover, Lemonidis et al. suggested that a Tn5801-like transposon, usually carrying the tet(M) gene, is frequently found in vanA-type pstS-null strains [22]. However, neither the tet(M) gene nor the Tn5801 transposon was identified in the 700,907 and 741,160 genomes.
Analysis of the vanA clusterIn order to investigate the genetic basis of vancomycin susceptibility of the 8 isolates, the nucleotide sequence of the Tn1546 transposon has been thoroughly studied and compared to the prototype Tn1546 (GenBank accession no. M97297). WGS analysis revealed that the transposase and resolvase genes were missing, and two types of vanA cluster have been found (Table 2 and Fig. 1): (i) the first type, found in 7 isolates, showed an intact vanH/vanA/vanX cluster, the lack of vanR gene and the deletion of vanS gene and (ii) the second type, found in the 741,160 strain, was completely devoid of both vanR and vanS genes and showed a deletion of the vanH gene.
Fig. 1
Linear map of the Tn1546 prototype of E. faecium BM4147 (accession no. M97297.1) in comparison with defective Tn1546 transposons of VVEIta-S and VVEIta-R isolates using Easyfig tool (https://mjsull.github.io/Easyfig/). The van gene cluster is shown in orange and the truncated vanH is indicated as ΔvanH. The positions and transcriptional direction of the ORFs are represented with arrows. Some antibiotic resistance determinants and relevant genes described in this study are shown
The first type of vanA cluster showed the insertion of the IS1216 element at the 3′ end of the vanS gene. This insertion caused the deletion of a 5789 bp region of the Tn1546 wildtype including the vanR gene, a large region of vanS (only 14 bp at the 3′ end was maintained), the resolvase and the Tn1546 transposase. It is well-known that IS elements are highly mobile and can cause Tn1546 structural alterations [35].
vanR and vanS are part of a two-component signal transduction system that controls the expression of the vanH/vanA/vanX cluster mediating vancomycin resistance. Several papers reported that the total absence or modification of this system could be responsible for a vanA genotype/vancomycin-susceptible phenotype, as the transcription of the genes required for resistance cannot be regulated [6, 11, 13]. Furthermore, six out of seven isolates had an ISEfm1 transposase between vanX and vanY genes.
The second type of vanA cluster found in the 741,160 strain had no vanR and vanS genes and also showed a deletion of 544 bp at the 5′-end of the vanH gene (only 425/969 bp of the vanH gene has been maintained) due to the insertion of an IS1216 element. This IS1216 element was present in the vanA-plasmid of the 741,160 strain in multiple copies and their transposition could have contributed to the deletion of the vanH. Moreover, also an ISEfm1 element between vanX and vanY genes was detected.
It is well-known that a complete vanH/vanA/vanX operon is required for the development of a vancomycin-resistant phenotype [6]; thus, the presence of a truncated vanH gene could contribute to vancomycin susceptibility. For this reason, the 741,160 strain was not considered a VVEIta-S.
VVEIta-S resistance phenotype reversion and its stabilityWe selected the VVEIta-S 700907 isolate (the representative of the 7 strains with the type 1 vanA cluster) to verify its ability to revert to the vancomycin-resistant phenotype under laboratory conditions. We obtained adaptive mutants by plating the strain in agar supplemented with increasing concentrations of vancomycin. Mutants were able to evolve to full vancomycin resistance after a week, showing a vancomycin MIC of > 128 mg/L.
The 700,907 mutant (VVEIta-R) was subjected to WGS and compared to the parental VVEIta-S strain by BLASTN analysis. The mutant genome disclosed a 44 bp deletion (ACACGGTAGACTTAATACCGTAAGAACGAGCCGTTTTCGTTCTT) in the promoter region of vanH/vanA/vanX cluster causing its constitutive expression, as previously described (Fig. 1) [6, 11]. The VVEIta-R strain showed unvaried high levels of resistance to vancomycin even after induction, confirming the constitutive expression of the vanH/vanA/vanX cluster. Wagner et al. suggested that an alternative promoter conveys vanH/vanA/vanX expression independently of the vanR activator [11]. Indeed, upstream of the vanH gene, we found the same consensus sequences of P2 and P3 promoters previously described by Wagner et al. [11]. Genome comparisons revealed no other relevant alterations that could be associated to the phenotypic differences between these isogenic strains.
We further investigated the stability of the vancomycin resistance phenotype of the VVEIta-R under non-selective conditions (on antibiotic-free) for 15 days. After 5 days of passages, the revertant isolates were still resistant (MIC, > 128 mg/L); however, the resistance phenotype was unstable; since at the 11th day of growth in agar without vancomycin, all strains showed a reduced level of resistance (MIC, 8 mg/L) despite the vanA gene was still detected. These findings are consistent with the data of Wagner et al. which highlighted a temporal progression of most VVE-R isolates towards vancomycin susceptibility [6] in the absence of exposure to vancomycin.
In order to determine the genetic basis associated with the reduction of vancomycin resistance, a selected mutant strain (named VVEIta-R1) was subjected to WGS. Comparing the region upstream of the vanH gene of the VVEIta-R mutant highly resistant to vancomycin (MIC, > 128 mg/L) with that of the VVEIta-R1 mutant (MIC, 8 mg/L), we found only one point mutation (C to A) located at the beginning of the truncated vanS gene.
Plasmid location of vanA clusterWGS analysis of the seven VVEIta-S and the E. faecium 741,160 revealed that the vanA cluster was always localized on plasmids of size range 42–56 kb (G + C content 35%), all belonging to RepA_N replicon type (Table 2). Plasmids did not carry any antibiotic-resistance genes other than the vanA gene cluster. The relevant ORFs of the eight vanA plasmids are indicated in tables S2 to S9.
BLASTN analysis revealed that the 8 vanA plasmids were 99.36%–99.97% (coverage range 91–100%) identical to each other and overall showed the best coverage and nucleotide identity (33–37% and 98–99%, respectively) with the 83.6 kb plasmid (accession no. CP092571.1) of the Enterococcus faecium VRE-WC072. The high nucleotide identity of these plasmids harbored by the 8 different strains isolated from different wards suggested an intra-species spread of the same vanA plasmid among the hospital circulating enterococci.
Moreover, all eight vanA plasmids showed a DNA identity of 100% (coverage 48%) with the 39 kb pS177 plasmid (GenBank accession no. NC_014959) of the E. faecium strain from the USA, which typically carried this defective vanA gene cluster (Fig. 2).
Fig. 2
Circular maps of the vanA-containing plasmids of the 8 sequenced VVEIta-S in comparison with pS177 plasmid using BRIG software. Plasmids and transposons included in the analysis were as follows: (inner to outer circles) pS177 of E. faecium S177 (accession no. HQ115078), pEfm741160-vanA of E. faecium 741,160 (accession no. OR234015), pEfm731980-vanA of E. faecium 731,980 (accession no. OR234011), pEfm732558-vanA of E. faecium 732,558 (accession no. OR251469), pEfm733387-vanA of E. faecium 733,387 (accession no. OR234012), pEfm735902-vanA of E. faecium 735,902 (accession no. OR298096), pEfm742783-vanA of E. faecium 742,783 (accession no. OR251470), pEfm755686-vanA of E. faecium 755,686 (accession no. OR262468), and pEfm700907-vanA of E. faecium 700,907 (accession no. OR208591). Red arrows indicate the position and orientation of the genes of the pEfm700907-vanA used as reference; some antibiotic resistance determinants and relevant genes described in this study are shown
In VVE isolates, the vanA gene cluster has been described also on a conjugative plasmid [9, 10, 14]. Despite several attempts, our strains were unable to transfer the vanA plasmid to the E. faecium 64/3 recipient at frequencies detectable under laboratory conditions. However, the high identity of plasmids carried by the eight different strains demonstrates that the dissemination of these vanA plasmids could occur by co-resident plasmids mediating the transfer.
The copy number of vanA plasmid was measured in the VVEIta-S 700907 strain and in its relative mutants 700,907 VVEIta-R and VVEIta-R1. Both mutants harbored a higher copy number of the vanA plasmid compared with the VVEIta-S 700907 parental strain, respectively, with a 17 ± onefold increase in mutant 700,907 VVEIta-R1 and 40 ± onefold in mutant 700,907 VVEIta-R consistently to their MIC values as previously reported by Wagner et al. [11].
留言 (0)