
記住我


Chronic liver disease and cirrhosis is the 12th overall leading cause of death in the United States (1). Chronic liver disease can be caused by infectious agents, alcohol, and fatty liver disease. In this study, patients with hepatitis C virus (HCV)-induced liver disease were used as a model to study the progression of chronic liver disease. Although infected with the same virus, only 20%–25% of patients infected with HCV progress to cirrhosis over 25–30 years (2). The exact mechanisms of disease progression and reasons for variability in progression remain to be elucidated.
The liver is responsible for a wide variety of physiologic processes including regulation of energy homeostasis. It plays a major role in protein and amino acid (AA) metabolism and uses AAs for energy and disposal of nitrogenous waste (3). As liver disease progresses, alterations in whole-body protein homeostasis occur contributing to the development of sarcopenia (4). Sarcopenia is present in 40%–70% of patients with cirrhosis (5) and is a significant predictor of prognosis in patients. The mortality risk is more than 2-fold higher in patients with sarcopenia compared with non-sarcopenic patients. Sarcopenia is an independent predictor of mortality when compared with other measures of liver dysfunction such as Child-Pugh or Model for End-Stage Liver Disease (6). While it is known to be a prognostic factor, the pathogenesis of sarcopenia is incompletely understood. It is known that one of the underlying mechanisms to sarcopenia is anabolic resistance to stimuli such as branched-chain AAs (BCAAs) (7). By understanding the pathogenesis of sarcopenia in liver disease specifically, it may become possible to use noninvasive markers in the evaluation of the progression of disease and targets for possible treatment.
We used a standard-meal challenge to create a stress test for patients with compensated liver disease. The increased portal pressure after a meal, as well as increased metabolites, gave us the opportunity to see how both cirrhotic and non-cirrhotic livers can respond to an increased metabolic and pressure load. In patients with progressive, decompensated liver disease (Child-Pugh Score ≥10), there is a known increase in aromatic AAs (AAAs) and decrease in BCAAs (8). We wanted to evaluate these AAs in patients whose disease had not yet progressed and still have early compensated disease (Child-Pugh Score <7). The standard meal would stress the compensated liver both in terms of increased blood flow to the liver and by increasing metabolic demands. Studying the dynamic response to this stress may help elucidate mechanisms that reflect and are related to decreased reserve in early liver disease.
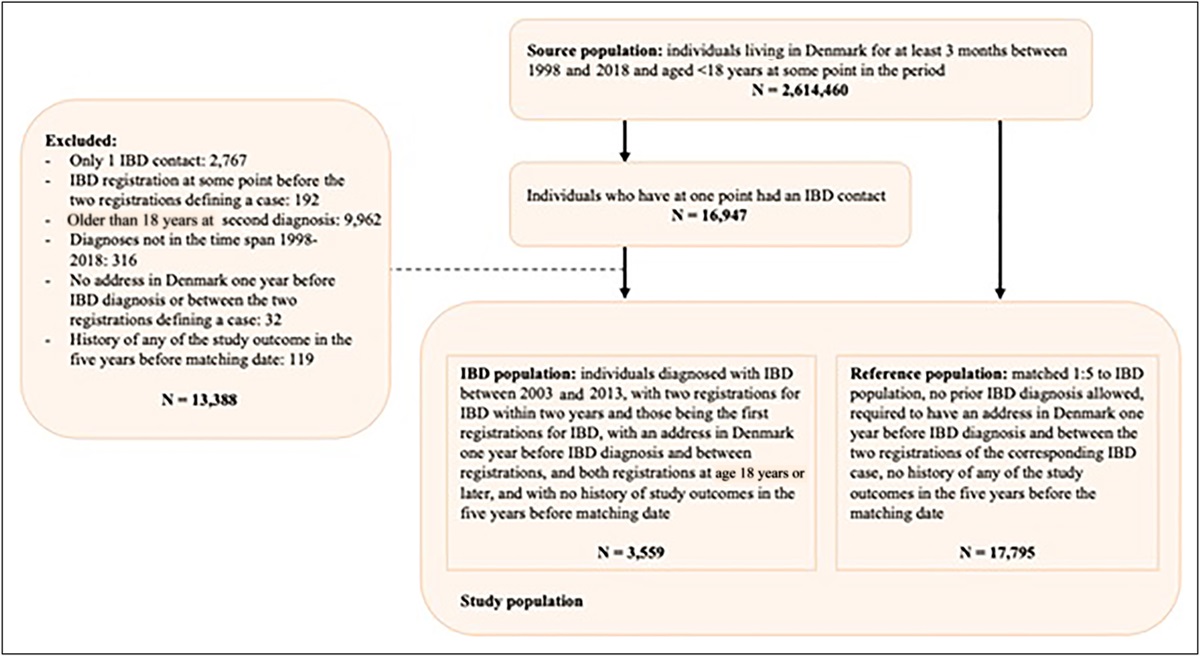
METHODS PatientsAs previously described, 30 patients were evaluated for chronic HCV-associated liver disease after exclusion of other forms of chronic liver disease at the National Institutes of Health Clinical Center (NCT02400216) (9). These patients were recruited as part of a broader study where patients were treated for HCV (9). All patients provided signed informed consent for participation in the Institutional Review Board-approved protocol. All HCV genotypes were included, and all patients had a Child-Pugh Score ≤6. Patients with hepatocellular carcinoma were excluded. Patients with an Ishak Score of 1–4 were considered non-cirrhotic, and patients with an Ishak Score of 5–6 were considered to have cirrhosis (C).
Sample collectionLiver biopsy and portal pressure measurements were obtained by percutaneous ultrasound-guided transhepatic puncture of an intrahepatic portal vein branch by interventional radiology. A transhepatic approach was selected in these patients because this procedure was part of a broader study which included sampling from the portal vein. Liver biopsy tissue was scored in a blinded manner by a single liver pathologist (D.E.K.) to obtain Ishak Fibrosis and Hepatic Activity Index scores for fibrosis and inflammation, respectively (10,11).
RNA-seq analysis of liver tissueLiver tissue samples (10 mg) were snap-frozen and stored at −80°C. Total RNA extraction was performed on the collected liver biopsies. To minimize the batch effect, samples were coded and processed in a blinded manner. This was done using TriZol (Cat No: 15596026) and Qiagen RNA Extraction Kit (Cat No: 74104). A poly-A selection was performed on the total RNA samples using the NEBNext Poly(A) Selection kit (Cat No: E7490S). The RNA library was prepared using the poly-A-selected RNA and ScriptSeq RNA Library Preparation Kit (Cat No: SSV21106). All cDNA libraries were quantified using the KAPA Biosystems Illumina qPCR Kit (Roche Cat No: 07960140001), normalized, and submitted to the NIDDK Genomics Core for Illumina HiSeq 4000 sequencing. Raw sequencing files were aligned to the Homo sapiens hg38 reference genome using STAR in Partek Flow (version 10.0) (12). Sequences were filtered using a cutoff threshold of <40 million reads with Phred scores <30 for all 50 bp sequenced. Two patients failed quality control and were excluded from analysis involving liver transcriptome data (n = 27).
The DESeq2 R package (version 1.32.0) was used to perform differential gene expression analysis, where the non-cirrhotic patient data were used as the reference. The analysis provided statistical data for 19,960 hepatic genes (13). The log2 ratio of the mean of normalized counts was overlaid on Kyoto Encyclopedia of Genes and Genomes pathways related to AA metabolism using the Pathview R package (version 1.32.0) (14). Data analysis and figure generation was conducted in R (version 4.1.0) (15) using the tidyverse collection of packages (version 1.3.1) (16).
Clinical markersBiochemical assays were performed on a cobas c 501 system (Roche Diagnostics) to measure alanine aminotransferase, aspartate aminotransferase, albumin, and total and direct bilirubin. Platelet counts were also measured. Complete blood counts were drawn in 3 mL K2 ethylenediamine tetraacetic acid tubes (Ref 367856; Becton, Dickinson and Company, Franklin Lakes, NJ) and measured on the Sysmex system. Quantifiable HCV RNA was measured by reverse-transcription polymerase chain reaction.
HepQuant SHUNT fractionFlow-dependent clearance of cholate was measured from both portal and systemic circulations to quantify the HepQuant SHUNT. This quantifies hepatic acinar function and effective sinusoidal perfusion. This test is able to measure hepatic filtration rates, portal-systemic spillover of d4-cholate (SHUNT%), d4-cholate at 60 minutes (STAT), and a Disease Severity Index (17).
Standard mealThe patients fasted overnight and were then given a standard-meal shake. This shake consisted of 315 g Lactaid whole milk, 190 g Lactaid vanilla ice cream, 10 g vanilla extract, 60 g granulated sugar, and 40 g Beneprotein powder. The nutritional values for each serving of the shake were 986.8 kcal, 51 g protein, 33.3 g fat, and 124.3 g carbohydrates. Five milliliters of blood was drawn from the antecubital vein into an ethylenediamine tetraacetic acid tube 60 and 30 minutes before the meal; with the meal; as well as 30, 60, 90, 120, 180, and 240 minutes after the meal. All serum samples were processed by centrifugation at 1000 rpm for 10 minutes within 4 hours of being drawn and subsequently stored as plasma at −80°C until analysis.
AA concentrationQuantitative analysis of AAs was performed at Mayo Clinic Laboratories using liquid chromatography-tandem mass spectrometry (Test ID: AAQP). According to the protocol, AAs present in plasma were labeled with a TRAQ Reagent 121. Samples were dried and reconstituted with a TRAQ Reagent 113-labeled Standard Mix. The AAs were separated and detected by liquid chromatography-tandem mass spectrometry, and concentrations were established by comparison of their ion intensity (121-labeled AAs) with that of their respective internal standards (113-labeled AAs).
Statistical analysisStatistical analysis was performed using Graphpad Prism 8.0 and R software. All nonpaired statistical tests were performed using 2-sided Mann-Whitney (MW) U test; data distribution was assumed to be normal but not formally tested.
RESULTS Patient demographicsOf the 30 patients chronically infected with HCV, 1 was excluded with cancer and the remaining 29 were evaluated in this study. Thirteen patients had cirrhosis, and 16 were non-cirrhotic. All patients with cirrhosis had a Child-Pugh Score A (compensated liver disease), and there was no significant difference in HCV loads between non-cirrhotic patients and patients with cirrhosis (Table 1). On average, all patients had increased alanine aminotransferase and aspartate aminotransferase concentrations, but total bilirubin, albumin, and platelets were within the reference intervals (Table 1). There was no significant difference between body mass index in non-cirrhotic patients and those with cirrhosis (Table 1). There was an association between sex and cirrhosis (χ2 test, P = 0.005), where male patients were more frequently in the non-cirrhotic group than female patients.
Table 1. - Baseline characteristics of patients Non-cirrhotic (N = 16) With cirrhosis (N = 13) P value Age (yr) 55.1 58.3 NS (0.97) Sex (male), % (n) 87.5 (14) 30.8 (5) 0.005a Log (HCV RNA) 6.6 6.9 NS BMI (kg/m2) 26.5 29.2 NS HbA1c (%) 5.5 5.5 NS ALT (IU/L) 105.9 112.2 NS AST (IU/L) 76.4 93.3 NS Total bilirubin (mg/dL) 0.6 0.8 0.01 Albumin (g/dL) 4.3 4.1 0.0 Platelets (×109/L) 182.0 138.2 0.01 Ishak Fibrosis 2.1 5.8 <0.0001 Hepatic Activity Index 7.9 8.5 NS Direct portal pressure 13.8 23.1 0.0012Values are means. P values were calculated with Mann-Whitney test.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; BMI, body mass index; HCV, hepatitis C virus; HbA1c, hemoglobin A1c; NS, not significant.
aThis value was calculated with χ2 test of independence.
While looking at the entire transcriptome, in patients with cirrhosis, there was increased downregulation in the AA process (9). When the log fold changes between cirrhotic and non-cirrhotic hepatic gene expression data were overlaid on Kyoto Encyclopedia of Genes and Genomes pathways relating to AA metabolism, the pathways related to AAA metabolism showed more downregulation in patients with cirrhosis compared with non-cirrhotic patients than the pathways related to BCAA metabolism. This is demonstrated by the presence of more downregulated (blue) gene nodes in patients with cirrhosis on the tyrosine metabolism pathway, which displays both tyrosine and phenylalanine metabolism (Figure 1a) as well as the tryptophan pathway (Figure 1b) than the number of gray gene nodes on the valine, leucine, and isoleucine (BCAAs) degradation pathway (Figure 2). The TAT1 transporter, responsible for the basolateral efflux of AAAs (18), was downregulated in patients with cirrhosis.
AAA metabolism is decreased in cirrhotic patient transcriptome when compared with non-cirrhotic patient transcriptome. (a) Tyrosine and phenylalanine metabolism. (b) Tryptophan metabolism. Blue, gray, red: see scale for corresponding log fold change value. White: gene was not measured in hepatic transcriptome. AAA, aromatic amino acid.


 Figure 2.:
Figure 2.: Transcriptome BCAA degradation. There is less fold change in BCAA degradation when cirrhotic patient transcriptome is compared with non-cirrhotic patient transcriptome. Blue, gray, red: see scale for corresponding log fold change value. White: gene was not measured in hepatic transcriptome. BCAA, branched-chain amino acid.
Baseline AA concentrations in compensated liver diseaseAAA (phenylalanine, tryptophan, and tyrosine) levels and BCAA (isoleucine, leucine, and valine) levels were summed at each time point. Because the main pathways of AAA metabolism and BCAA metabolism are common, single AAs were not analyzed. When evaluated at baseline, there was no significant difference in AAA concentrations between patients with cirrhosis and non-cirrhotic patients (Figure 3a). Median AAA concentration at baseline was 255 nmol/mL of patients with cirrhosis (interquartile range [IQR] 63 nmol/mL) while the median of non-cirrhotic patients at baseline was 251 nmol/mL (IQR 44 nmol/mL). There was also no significant difference in BCAA concentration at baseline between patients with cirrhosis and non-cirrhotic patients (Figure 3b). The median BCAA concentration at baseline of patients with cirrhosis was 506 nmol/mL (IQR 97 nmol/mL) and 533 nmol/mL (IQR 144.8 nmol/mL) for non-cirrhotic patients.
 Figure 3.:
Figure 3.: (a, b) Amino acid concentrations at baseline in patients with cirrhosis and non-cirrhotic patients. There was no significant difference in AAAs or BCAAs at baseline between patients with cirrhosis and non-cirrhotic patients. Solid circles represent non-cirrhotic patients. Outlined circles represent patients with cirrhosis . Error bars represent median with range. AAA, aromatic amino acid; BCAA, branched-chain amino acid.
Dynamic AA concentrationsAlthough no difference was seen at baseline, 30 minutes after the standard meal, patients with cirrhosis had a significantly higher level of AAAs than non-cirrhotic patients (MW, P = 0.039). This significant difference in AAAs was found at all subsequent time points (60, 90, 120, 180, 240; MW: 0.016, 0.0014, 0.0041; 0.0003, 0.0021). There was no significant difference between patients with cirrhosis and non-cirrhotic patients in BCAAs at any time points after the standard meal was given (Figure 4a, b).
 Figure 4.:
Figure 4.: (a, b) Dynamic amino acid concentrations after a standard meal. AAAs were significantly different between patients with cirrhosis and non-cirrhotic patients at all time points after the meal. BY contrast, there was no significant difference between BCAAs in patients with cirrhosis and non-cirrhotic patients after the meal. Solid circles represent non-cirrhotic patients. Outlined circles represent patients with cirrhosis. Error bars represent median with range. ****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05. AAA, aromatic amino acid; BCAA, branched-chain amino acid.
Peak changeThe peak change was calculated by subtracting the highest postprandial concentration of summed AAs from the baseline value. This represents the delta between baseline AA concentrations and the point of maximum AA infiltration through the liver. Patients with cirrhosis had a significantly higher peak change of AAAs compared with non-cirrhotic patients (MW, P = 0.0073). Patients with cirrhosis had a median delta AAA concentration of 161 nmol/mL (IQR 156 nmol/mL) while non-cirrhotic patients had a median delta AAA concentration of 114 nmol/mL (IQR 45.8 nmol/mL). There was no difference seen in peak change of BCAAs between patients with cirrhosis and non-cirrhotic patients (Figure 5a, b). Patients with cirrhosis had a median delta BCAA concentration of 581 nmol/mL (IQR 320 nmol/mL) while non-cirrhotic patients had a median delta BCAA concentration of 511 nmol/mL (IQR 228.3 nmol/mL). Considering patients with cirrhosis have decreased transcription of genes in the AAA metabolism pathway, these dynamic AA differences suggest patients with cirrhosis to have less ability to cope with increased AAA load after a meal.
 Figure 5.:
Figure 5.: (a, b) Peak change in amino acid concentrations after a standard meal. The peak change in AAAs after the meal was significantly higher in patients with cirrhosis compared with non-cirrhotic patients. There was no difference seen in the peak change of BCAAs between patients with cirrhosis and non-cirrhotic patients. Solid circles represent non-cirrhotic patients. Outlined circles represent patients with cirrhosis. Error bars represent median with range. AAA, aromatic amino acid; BCAA, branched-chain amino acid.
Blood flow bypassing the liver by shunting correlates with only AAAs, not BCAAsThe HepQuant SHUNT fraction was measured at baseline by first pass hepatic extraction of cholate and represents the percentage of portal blood that bypasses the liver parenchyma through intrahepatic or extrahepatic shunts. The HepQuant SHUNT fraction was significantly elevated in patients with cirrhosis compared with non-cirrhotic patients (MW, P = 0.0007) (Figure 6). The median HepQuant SHUNT fraction in patients with cirrhosis was 34.7% (IQR 12.1%) while non-cirrhotic patients had an average of 24.9% (IQR 6.2%). The HepQuant SHUNT fraction positively correlated with the levels of AAAs at baseline as well as at all time points after a standard meal (Table 2). Interestingly, there was no relationship found between BCAAs at any time point with the HepQuant SHUNT fraction. This suggests that increased AAAs in patients with cirrhosis is more likely due to decreased transcription of enzymes required in metabolism rather than shunting AAs away from the liver.
 Figure 6.:
Figure 6.: Blood shunting around the liver parenchyma at baseline. HepQuant SHUNT fraction was significantly higher in patients with cirrhosis compared with non-cirrhotic patients. Solid circles represent non-cirrhotic patients. Outlined circles represent patients with cirrhosis. Error bars represent median with range.
Table 2. - Blood shunting around the liver parenchyma correlates with AAA concentration before and after a standardized meal Baseline 30 60 90 120 180 240 BCAA NS NS NS NS NS NS NS AAA *** (0.60) ** (0.52) *** (0.61) *** (0.64) ** (0.55) *** (0.62) ** (0.57)The HepQuant SHUNT fraction positively correlated with the AAA levels at baseline and postprandially but was not correlated with the BCAA concentrations. Value represents Spearman r. Time is measured in minutes after the standard meal.
AAA, aromatic amino acid; BCAA, branched-chain amino acid; NS, not significant.
****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05.
It is known that the ratio of AAAs to BCAAs is high in decompensated liver disease (8). Decompensated liver disease occurs when patients begin to experience symptoms associated with portal hypertension, including ascites, spontaneous bacterial peritonitis, variceal bleeding, and hepatic encephalopathy (19). In our study, we were able to create a stress test for the liver by giving patients with compensated liver disease a standard meal. In healthy individuals, portal blood flow increases by approximately 59% after a meal (20), and by giving our patients a standard meal, we are able to create a temporary state of increased portal pressure and increased metabolic load. We then measured AAs postprandially and observed a significantly larger increase in the peripheral concentration of AAAs in patients with compensated cirrhosis (Ishak 5–6) when compared with non-cirrhotic patients (Ishak 1–4). Higher AAA concentration at baseline has been shown in decompensated liver disease, but it has not yet been seen in patients with compensated disease. After a standardized meal, in the setting of increased mechanical and metabolic stress to the liver, we see an elevation of AAAs in the compensated cirrhotic cohort. Interestingly, there was no significant difference between BCAA concentrations in patients with cirrhosis and non-cirrhotic patients at baseline or postprandially.
The increase in AAA levels is consistent with observations in the hepatic transcriptome. The hepatic transcriptome in patients with cirrhosis showed greater downregulation of genes associated with AAA degradation compared with those associated with degradation of BCAAs. This could imply that in a non-stressed state, such as fasting, compensated cirrhotic livers can synthesize enough enzymes to metabolize the baseline amount of AAAs presenting to the liver. However, with increased influx of AAAs into a compensated cirrhotic liver, there is already enough underlying dysfunction such that the liver is unable to increase transcription for AAAs the same way non-cirrhotic patients can. This results in significantly increased AAA concentration in serum of patients with cirrhosis compared with non-cirrhotic patients after a meal. This is also the possible reason a difference at baseline is not seen. It is intriguing to speculate that the decreased ability to degrade AAAs is not dysfunctional per se, but rather the result of active inhibition of degradation to allow for availability of AAAs for possibly more important functions elsewhere in the body. It is known that during early HCV infection, there is metabolic reprograming favoring increased AA concentrations in response to increased metabolic load. It has also been seen in cell culture that there is an increase in uptake of AAs from medium in response to increased anabolic and energy demands (21). Interestingly, in patients with cirrhosis, there was also downregulation of the basolateral AAA efflux transporter, TAT1. This transporter has been found on the epithelial cells of the small intestine and kidney and is known to play a role in homeostasis of AAs (22). Although requiring validation, this finding does highlight a possible connection between TATA1 receptors on liver epithelia and AA homeostasis in cirrhotic liver disease.
Decreased BCAA concentrations are believed to be due to, in part, elevated levels of ammonia caused by increased portosystemic shunting during liver disease (23). The liver is responsible for metabolism of ammonia through the urea cycle, and when ammonia is shunted past the liver, it is not metabolized, leading to increased ammonia levels. This increased ammonia stimulates glutamine synthesis, which reduces the glutamate store and, therefore, indirectly BCAAs (24). Using the cholate shunt, we were able to calculate the percentage of blood being shunted away from the liver. Interestingly, this value was associated with AAAs at baseline and postprandially but not with BCAAs at any time point. This shows that portosystemic shunting is related to the increased AAA concentrations seen in compensated liver disease. Because there is no association between BCAAs and shunting, it is less likely that portosystemic shunting plays a significant role in BCAA concentrations in compensated liver disease. It may only play a role in more severe, uncompensated disease or the change in BCAA concentrations may be related to other mechanisms.
Shunting blood, and thus excluding the liver, has also been postulated as a contributing factor to the increased AAA concentration in decompensated liver disease (8). AAAs are metabolized by the liver, and decreased hepatic function will lead to increased levels of AAAs. Our study showed a significant association between the HepQuant SHUNT fraction and AAA concentration, which indicates that there is a relationship between AAAs, not BCAAs, and liver disease severity before decompensation. Most studies on the effect of shunting in liver disease and associated changes in AA levels have been performed in patients who have had the TIPS procedure (25,26). TIPS could be an independent factor related to perturbations in AA levels. Our study yields insight into these changes in compensated patients and how these AA perturbances occur in cirrhosis before decompensation and hepatic encephalopathy.
Owing to derangements in liver function, patients with cirrhosis have insufficient glycogen stores. Their ability to generate energy after an overnight fast is comparable with that of a healthy individual who has fasted for 2–3 days (27). Therefore, patients with cirrhosis are in a chronic catabolic state. The resting energy expenditure of over 50% of patients with cirrhosis studied was raised more than 120% of the expected value (28). HCV-induced cirrhosis is a chronic inflammatory state. Patients not only lack energy stores because of decreased glycogen but they are also expending increased amounts of energy because of the inflammatory response amounted against HCV. It is believed that as a mechanism of keeping up with increased energy needs patients with cirrhosis to break down muscle for energy generation (28,29). Sarcopenia is a common complication of end-stage liver disease, leading to significant morbidity. It is a strong predictor of adverse clinical outcomes in patients with cirrhosis (29).
All AAAs (tyrosine, phenylalanine, and tryptophan) are ketogenic AAs and metabolized in the liver and adrenal glands. Thus, they can be metabolized to acetyl-CoA, incorporated in the TCA cycle, and play a role in energy production (26). In patients with cirrhosis, higher concentrations of AAAs were found, which is likely because of the decreased metabolism by the liver. This decreases acetyl-Co availability to reduce pyruvate and incorporate citrate into the TCA cycle. With decreased acetyl-CoA from AAAs, which are normally metabolized in the liver, other reservoirs of energy will be expended. This might be one of the mechanisms in which sarcopenia develops in liver disease. The lean muscle mass will be broken down for ketogenic AAs that can be expended for energy independent of the liver. This too may be a mechanism to maintain the availability of AAAs for other essential functions.
BCAAs are metabolized by both ketogenic and glucogenic pathways in the muscle, adipose tissue, kidneys, and brain. The ketogenic BCAAs (leucine and isoleucine) are first metabolized by BCAA aminotransferase in the skeletal muscle and then in the liver by alpha-keto acid dehydrogenase in the mitochondrial inner membrane yielding acetyl-CoA. The glucogenic BCAAs (isoleucine and valine) are catabolized in a similar mechanism, but yield succinyl-CoA, which can continue through gluconeogenesis. Isoleucine is both glucogenic and ketogenic. The BCAAs remain similar across both non-cirrhotic patients and patients with cirrhosis. In cirrhosis, impaired mitochondrial function has been shown to play a role in the impaired energy homeostasis. Therefore, we might not see a difference in BCAA levels during compensated cirrhosis because although decreased, patients are not yet sarcopenic and have maintained enough lean muscle function to complete the first step in degradation of the BCAAs despite the increase influx after a meal. As liver disease progresses and the muscle mass decreases, in some patients, there might be a tipping point where the residual muscle mass is unable to use enough BCAAs to maintain energy homeostasis and the patient will become decompensated. It would be after this tipping point that increased AAAs and decreased BCAAs can be seen at baseline.
These findings from a small sample size are associative and would need to be further studied in a controlled experimental setting. Our liver transcriptome was sequenced from a liver biopsy that was taken fasting; thus, information about hepatic transcription after the meal stress is limited. Obtaining a liver biopsy after a standard meal and comparing it with baseline would provide further information on the ability of patients with cirrhosis to adapt to increased metabolic needs. Our patients were infected with HCV, and this could potentially play a role in the affected AA concentrations in the meal study. Owing to direct-acting antiviral agents and the ability to eliminate HCV, it would also be interesting to repeat this study in patients after the clearance of HCV. It would also be thought-provoking to evaluate this in other etiologies of liver disease such as other viral hepatitis or metabolic dysfunction-associated steatotic liver disease. There was a significant difference in sex between the cirrhotic and non-cirrhotic groups, which may have played a role in AA concentration because of difference in gender lean muscle mass. Finally, it would have been very interesting to examine the transcriptome from a muscle biopsy obtained at the same time point. This would help better understand the liver-muscle axis in the progression of liver disease.
In summary, studying patients with compensated cirrhosis gives us insight into the progression of liver disease. Instead of focusing on the lack of BCAAs as playing a key role in the progression of liver disease and sarcopenia, it might be the AAAs that are playing a more significant role. It would be interesting to further investigate the biological role of AAAs in sarcopenia in liver disease to understand the mechanisms that differentiate AAAs from BCAAs. This would also bring light to the liver-muscle axis in the progression of liver disease. Measurement of AAA concentration might be a more sensitive test to indicate reserve of liver function in cirrhosis and could be used as an earlier, noninvasive test to evaluate hepatocyte metabolic function and ability to maintain energy homeostasis.
CONFLICTS OF INTERESTGuarantor of the article: Kareen L. Hill, MD.
Specific author contributions: All authors had a substantial contribution to this work. All authors provided approval for the final submitted version of the manuscript. Individual contributions are listed as follows: K.L.H.: design of the work; acquisition, analysis, interpretation of data; drafted the initial work and substantively revised it. J.A.H.: analysis and interpretation of data, substantively revised the work. R.O.A.: design of the work, acquisition and analysis. G.Y.Z.: acquisition, analysis, and interpretation of data. G.M.Q.: acquisition, analysis, and interpretation of data. E.T.: acquisition, analysis, and interpretation of data. G.T.E.: acquisition of data. S.M.H.: acquisition of data. M.B.: analysis of data. C.K.: acquisition of data. E.B.L.: acquisition of data. D.E.K.: acquisition of data. D.B.S.: acquisition of data. O.E.: design of the work; acquisition, analysis, and interpretation of data; substantively revised the work. T.H.: conception and design of the work; acquisition, analysis, and interpretation of data; drafted the initial work and substantively revised it.
Financial support: Financial support was provided by the intramural programs of the National Institute of Diabetes and Digestive and Kidney Diseases (DK054514), National Cancer Institute, and Clinical Center of the National Institutes of Health. In addition, the project was funded by an intramural NIH Bench to Bedside and Back Program Award: Mechanisms of microbial translocation in hepatitis C related liver disease 2014.
Potential competing interests: None to report.
Study Highlights
WHAT IS KNOWN ✓ In patients with progressive, decompensated liver disease (Child-Pugh Score ≥10), there is a known increase in aromatic amino acids (AAAs) and decrease in branched-chain amino acids (BCAAs). ✓ Sarcopenia is present in 40%–70% of patients with cirrhosis and is a significant predictor of prognosis in patients. ✓ A meal can stress the compensated liver both in terms of increased blood flow to the liver and by increasing metabolic demands. WHAT IS NEW HERE ✓ AAA elevation in patients with compensated cirrhosis seen after a standardized meal, which has only previously been seen in decompensated patients. ✓ In cirrhotic patients, elevation in AAAs correlated with increased downregulation of genes associated with AAA degradation but not those associated with degradation of BCAAs ✓ HepQuant SHUNT fraction correlating with AAA levels after a meal. ACKNOWLEDGMENTSWe thank the patients, staff, Michael W. Krause, Jim E. Balow, T. Jake Liang, Jay H. Hoofnagle, and the Institutional Review Board.
REFERENCES 1. Heron M. Deaths: Leading causes for 2009. Natl Vital Stat Rep 2012;61(7):1–94. 2. Lingala S, Ghany MG. Natural history of hepatitis C. Gastroenterol Clin North Am 2015;44(4):717–34. 3. Trefts E, Gannon M, Wasserman DH. The liver. Curr Biol 2017;27(21):R1147–51. 4. Ebadi M, Bhanji RA, Mazurak VC, et al. Sarcopenia in cirrhosis: From pathogenesis to interventions. J Gastroenterol 2019;54(10):845–59. 5. Kim G, Kang SH, Kim MY, et al. Prognostic value of sarcopenia in patients with liver cirrhosis: A systematic review and meta-analysis. PLoS One 2017;12(10):e0186990. 6. Montano-Loza AJ, Meza-Junco J, Prado CMM, et al. Muscle wasting is associated with mortality in patients with cirrhosis. Clin Gastroenterol Hepatol 2012;10(2):166–73.e1. 7. Kobayashi H. Amino acid nutrition in the prevention and treatment of sarcopenia [in Japanese]. Yakugaku Zasshi 2018;138(10):1277–83. 8. Holecek M. Ammonia and amino acid profiles in liver cirrhosis: Effects of variables leading to hepatic encephalopathy. Nutrition 2015;31(1):14–20. 9. Ali RO, Quinn GM, Umarova R, et al. Longitudinal multi-omics analyses of the gut-liver axis reveals metabolic dysregulation in hepatitis C infection and cirrhosis. Nat Microbiol 2023;8(1):12–27. 10. Ishak K, Baptista A, Bianchi L, et al. Histological grading and staging of chronic hepatitis. J Hepatol 1995;22(6):696–9. 11. Knodell RG, Ishak KG, Black WC, et al. Formulation and application of a numerical scoring system for assessing histological activity in asymptomatic chronic active hepatitis. Hepatology 1981;1(5):431–5. 12. Partek Flow. Version 10.0. Partek: Chesterfield, MO, 2020. 13. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014;15(12):550. 14. Luo W, Brouwer C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013;29(14):1830–1. 15. R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2021. 16. Wickham H, Averick M, Bryan J, et al. Welcome to the tidyverse. J Open Source Softw 2019;4(43):1686. 17. Wieland A, Etzion O, Ali RO, et al. HepQuant SHUNT detects portal hypertension in early stages of clinically compensated chronic liver disease. Clin Gastroenterol Hepatol 2022;20(4):e890–4. 18. Ramadan T, Camargo SM, Summa V, et al. Basolateral aromatic amino acid transporter TAT1 (Slc16a10) functions as an efflux pathway. J Cell Physiol 2006;206(3):771–9. 19. Poordad FF. Presentation and complications associated with cirrhosis of the liver. Curr Med Res Opin 2015;31(5):925–37. 20. Gaiani S, Bolondi L, Li Bassi S, et al. Effect of meal on portal hemodynamics in healthy humans and in patients with chronic liver disease. Hepatology 1989;9(6):815–9. 21. Roe B, Kensicki E, Mohney R, et al. Metabolomic profile of hepatitis C virus-infected hepatocytes. PLoS One 2011;6(8):e23641. 22. Bröer S. Amino acid transport across mammalian intestinal and renal epithelia. Physiol Rev 2008;88(1):249–86. 23. Katayama K. Zinc and protein metabolism in chronic liver diseases. Nutr Res 2020;74:1–9. 24. Holm E, Sedlaczek O, Grips E. Amino acid metabolism in liver disease. Curr Opin Clin Nutr Metab Care 1999;2(1):47–53. 25. Olde Damink SW, Jalan R, Redhead DN, et al. Interorgan ammonia and amino acid metabolism in metabolically stable patients with cirrhosis and a TIPSS. Hepatology 2002;36(5):1163–71. 26. Dejong CH, van de Poll MC, Soeters PB, et al. Aromatic amino acid metabolism during liver failure. J Nutr 2007;137(6 Suppl 1):1579S–85S; discussion 1597S–8S. 27. Owen OE, Trapp VE, Reichard GA Jr, et al. Nature and quantity of fuels consumed in patients with alcoholic cirrhosis. J Clin Invest 1983;72(5):1821–32. 28. Müller MJ, Lautz HU, Plogmann B, et al. Energy expenditure and substrate oxidation in patients with cirrhosis: The impact of cause, clinical staging and nutritional state. Hepatology 1992;15(5):782–94. 29. Dasarathy S, Merli M. Sarcopenia from mechanism to diagnosis and treatment in liver disease. J Hepatol 2016;65(6):1232–44.
留言 (0)