In our study, we phenotypically and genotypically analysed 82 MRSA strains isolated from patients with bloodstream infections in a central region of Slovenia in 2019–2022. This represents approximately one third of Slovenian MRSA isolates submitted to EARS-Net.
In our study, three SCCmec types (II, IV, V), eight ST types (ST1, ST5, ST22, ST45, ST97, ST225, ST398, ST2883), six CC complexes (CC1, CC5, CC22, CC45, CC97, CC398) and 16 spa types (t002, t003, t010, t011, t014, t022, t045, t127, t359, t790, t1227, t2770, t4336, t6890, t11581, t19843) were identified.
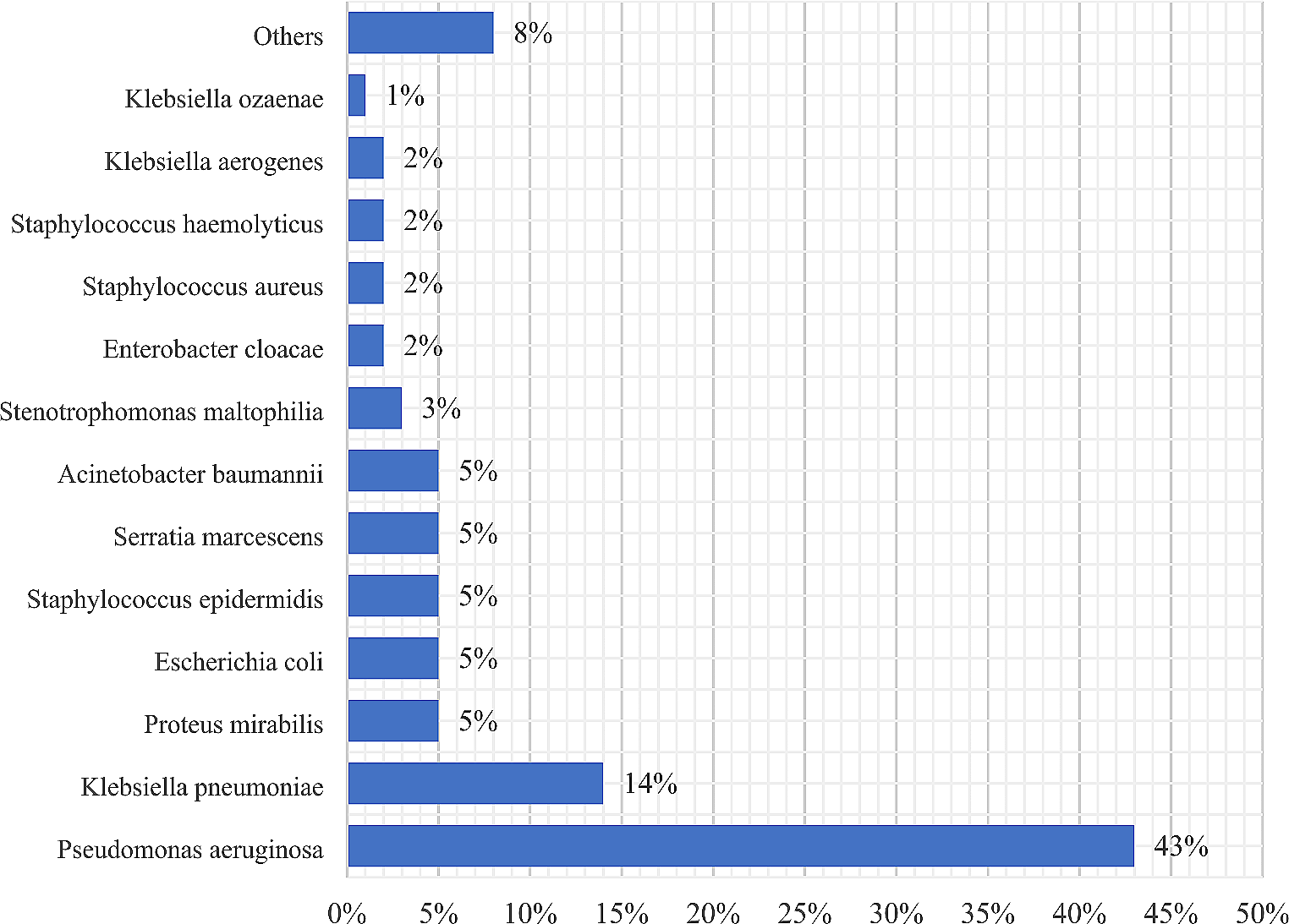
According to SKUOPZ data, MRSA isolates from clinical samples in Slovenia in 2017 were resistant to erythromycin, clindamycin, and ciprofloxacin in 75.2%, 74.3%, and 73%, respectively [10]. In our study we show that the majority of MRSA strains (n = 59) were also resistant to multiple antimicrobials (erythromycin, clindamycin, and ciprofloxacin) and belonged to ST225 (n = 36), ST2883 (n = 13), ST22 (n = 7), ST5 (n = 2) and ST97 (n = 1). MRSA strains (n = 20) that were susceptible to erythromycin, clindamycin and ciprofloxacin belonged to ST1 (n = 1), ST5 (n = 1), ST22 (n = 1), ST45 (n = 2), ST97 (n = 13), and ST398 (n = 2). The concordance of phenotype and genotype with the identified resistance gene was 97.8%. Discordant results for penicillin resistance could be due to incorrect reading of the edge of the inhibition zone as fuzzy or sharp according to EUCAST standards [18]. Discrepancies between phenotype and genotype resistance could either be due to the isolates having a nonexpressed resistance gene, plasmid loss, the isolates lacking the resistance gene or conferring other resistance mechanisms. All isolates of our most prevalent ST225-II and ST2883-II carried resistance genes against erythromycin and exhibited phenotypic resistance. Macrolide, ciprofloxacin, and mupirocin resistance phenotypes were described as biomarkers of epidemiological success in a recent study by Baede et al., whereas gentamicin resistance was associated with sporadicity [33]. These observations are consistent with our results.
CC5 (n = 53, 64.7%) was the predominant clone identified among MRSA strains and is the most prevalent clone causing HA-MRSA infections in the Western hemisphere [34]. In our study, CC5 included ST5, ST225 and ST2883.
ST225-II, also known as the Rhine Hesse MRSA clone [35] was identified in 37 isolates (45.1%). The most common spa type in our study that belonged to ST225-II, t003 (n = 26), was also one of the most prevalent spa types causing invasive MRSA infections in Europe [7]. According to the study by Kotnik-Kervokijan et al. [13], these spa types t003 have been found in Slovenia previously, so ST225-II clone is probably one of the endemic clones circulating in our country. In our study, all ST225-II strains carried inducible resistance by the erythromycin ermA gene and the aminoglycoside resistance gene ant(9)-Ia, as well as the enterotoxin genes seg, sei, sem, sen and seo.
ST2883-II was identified in 13 isolates (15.9%), twelve of which belonged to spa type t4336, and one to t19843. A novel ST2883 was identified among suspected CA-MRSA in Slovenia in a previous study in 2010 [12]. To our knowledge, no further data are available. All ST2883-II MRSA isolates were resistant to penicillin, erythromycin, clindamycin, and ciprofloxacin, carried lukED genes, enterotoxin genes (seg, sei, sem, sen, seo, seu), and haemolysin genes (alpha, beta, gamma). Alleles of sequence types ST225 and ST2883 only differ in a single nucleotide in the gene yqiL. Based on pairwise allele differences, some of the isolates belonging to ST225 are more closely related to ST2883 than other isolates belonging to the same ST. The two sequence types are highly similar; of the seven genes used for typing, there is a single nucleotide difference in the yqiL gene. ST2883 clone is a novel variant of ST225 and an important clone circulating in Slovenia that causes bloodstream infections.
ST5-II was detected in two isolates (2.4%, spa type t002) and is a pandemic strain also known as the New York/Japan clone [35]. In 2011, spa type t002 was the fourth most common spa type among invasive MRSA in Europe according to Grundmann et al. [7]. Both strains harboured the macrolide resistance gene ermA, aminoglycoside resistance gene aadD and the virulence-associated genes sed, seg, sei, sem, sen, seo, ser, and seu. We also detected one (1.3%, t010) MRSA strain belonging to ST5-IV, also known as the Paediatric clone [35]. The strain was resistant to penicillin and carried the blaZ gene, the haemolysin genes (alpha, beta, gamma), the lukED gene, and- the enterotoxin genes (sed, seg, sei, sej, sem, sen, seo, ser, and seu).
The second most frequent clone was CC97, ST97-IVc, which was identified in 15 isolates (18.3%). Of these, 13 belonged to spa type t359, one to spa type t2770 and one was non-typable. This clone is usually associated with livestock and rarely with human infections [36, 37]. Spa type t359 was previously confirmed in Slovenia among suspected human CA-MRSA isolates in 2014 and 2015 [11]. ST97-IV-t359 has also been found in Bosnia and Herzegovina and in a maternity hospital in Ireland [38, 39]. Although reports of ST97-IV clone are less common in Europe, it is considered an important pathogen in Slovenia. In our study, all ST97 MRSA isolates were resistant to penicillin and cefoxitin and carried the lukED gene. The ST97 isolates were negative for exfoliative toxin genes, toxic shock syndrome toxin genes and enterotoxins genes, similarly to isolates of other major LA-MRSA clones (i.e., CC398 and CC1) [2]. Two ST97 isolates were sak and scn genes negative, indicating a possible animal origin [1, 2].
The third most frequent clone CC22, ST22-IV was identified in eight isolates (9.8%), of which three belonged to spa type t022, four to t790 and one to t11581. ST22-IV is a pandemic strain also known as EMRSA-15 [35]. According to the study by Grundmann et al. the spa type t022 was the ninth most common spa type among invasive MRSA in Europe in 2011 [7]. All isolates belonging to ST22-IV in our study were resistant to penicillin, 25% to ciprofloxacin, erythromycin and clindamycin, and harboured haemolysin (alpha, beta, gamma) and enterotoxin genes (seg, sei, sem, sen, seo, and seu).
We have also identified other clones associated with LA-MRSA: two isolates belonged to ST1-IVa (2.4%, spa type t127) and two belonged to ST398-V (2.4%, spa type t011). All strains were resistant to penicillin, tetracycline, 25% to erythromycin and clindamycin. spa type t127 carried the lukED gene and the enterotoxin gene sek. The sak and the scn genes were negative in one strain, spa type t011, ST398, indicating a possible animal origin [1, 2]. None of the ST398-V-t011 strains had genes associated with virulence (enterotoxins, Pantone-Valentine leukocidin). According to our previous study among suspected CA-MRSA [11, 12] and the Kotnik-Kervokijan study [13], the ST398 clone has spread in the community and healthcare settings throughout the country. Currently, the number of severe infections caused by ST398 remains low in a central region of Slovenia. Larsen and colleagues reported an increased number of LA-MRSA ST398 bacteremias in Denmark, where most patients lived in rural areas but had no contact with livestock. The researchers predicted that the number of severe infections and deaths will increase if LA-MRSA ST398 is allowed to spread in the general population [39].
The final CC45 clone, ST45-IV was identified in two isolates (2.4%, spa type t6890) [35]. Both strains were resistant to penicillin and tetracycline, and they carried haemolysin (alpha, beta, gamma) and enterotoxin genes (sec, e.g., sei, sel, sem, sen, seo, and seu).
Surprisingly, we did not find any mecC positive clone, otherwise found in previous studies, in which mecC positive strains were detected both in asymptomatic carriers and also in clinical samples (sputum, blood culture) [11, 12, 14].
The virulence factors of S. aureus play an important role during pathogenesis [1,2,3]. Overall, in our study, all MRSA isolates harboured exotoxin genes (hla, hlb and hlg), adhesin (icaA) and enzymes (sspA, sspB) which may facilitate invasion and immune evasion. We also confirmed that over 50% of bloodstream MRSA isolates encode seg, sei, sem, sen, seo and / or seu [40]. Surprisingly, all isolates that belonged to ST97 had none of the enterotoxin genes, while two isolates that belonged to ST1 harboured only seh. According to the Bennett and Thomsen study the leucocidin toxin genes lukED are common across S. aureus lineages, with approximately 87% clinical isolates carrying lukED [41]. A similar percentage (85.4%) was found in our study, and only MRSA strains that belonged to ST22, ST45 and ST398 were lukED negative.
The main limitation of our study is the incomplete access to medical data and data on possible patient transfers between hospitals. Therefore, we could not investigate possible transmission events or investigate correlation over time. Additionally, two isolates in our study were non-typeable by spa typing. Due to limitations of short-read sequencing when it comes to repetitive regions, as well as possible low sequencing coverage in this region, spa types could not be reliably determined, because multiple spa repeats were assembled, or the 5’ or 3’ signatures were missing. Spa types were considered reliable when only one repeat sequence was recognised and the 5’ and 3’ signatures were found in the correct positions, as was the case for other isolates. For further studies, long-read sequencing could be employed to resolve these limitations. Like other epidemiologic studies, our study is also limited by the selected isolates.


留言 (0)