
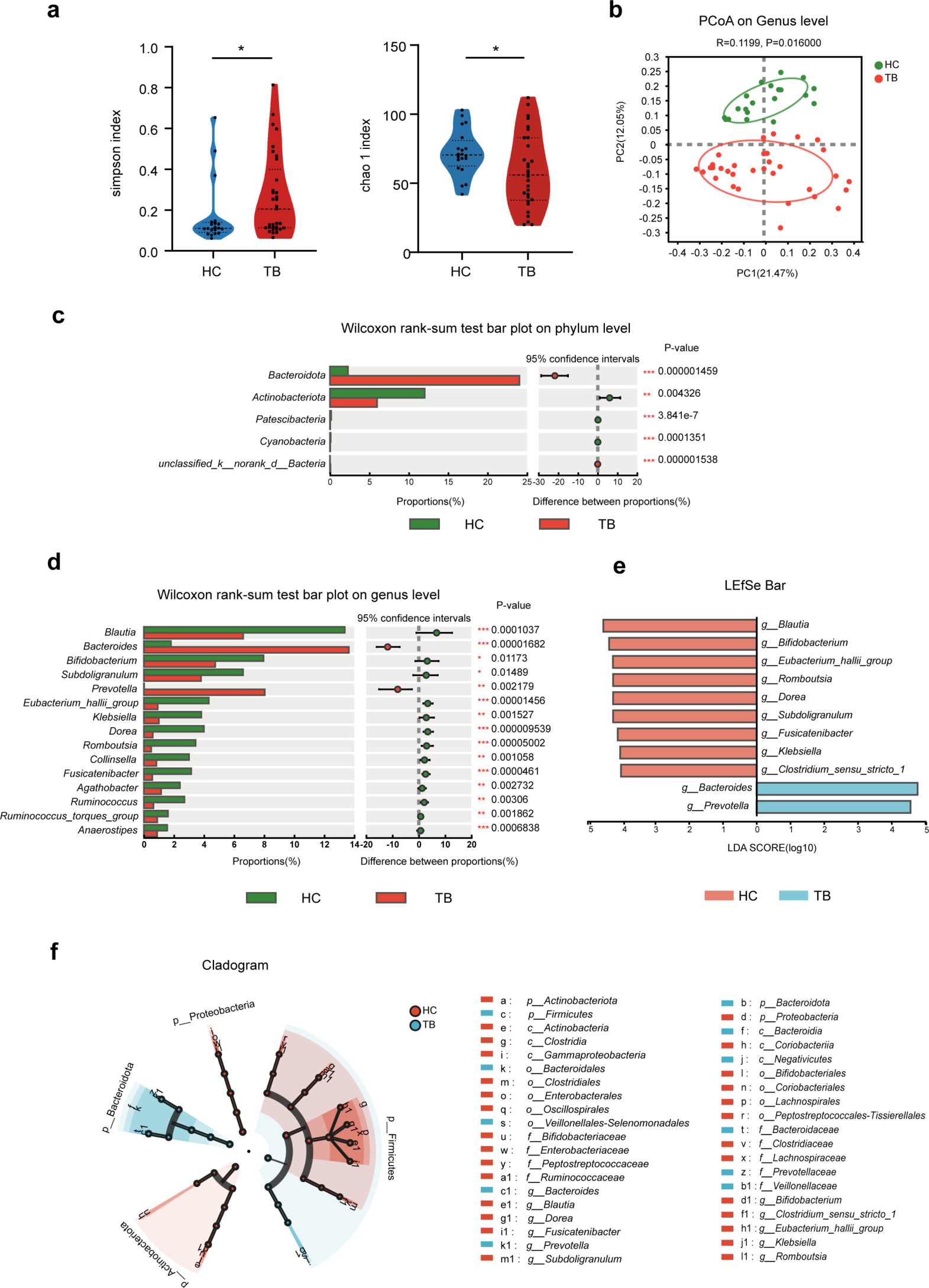
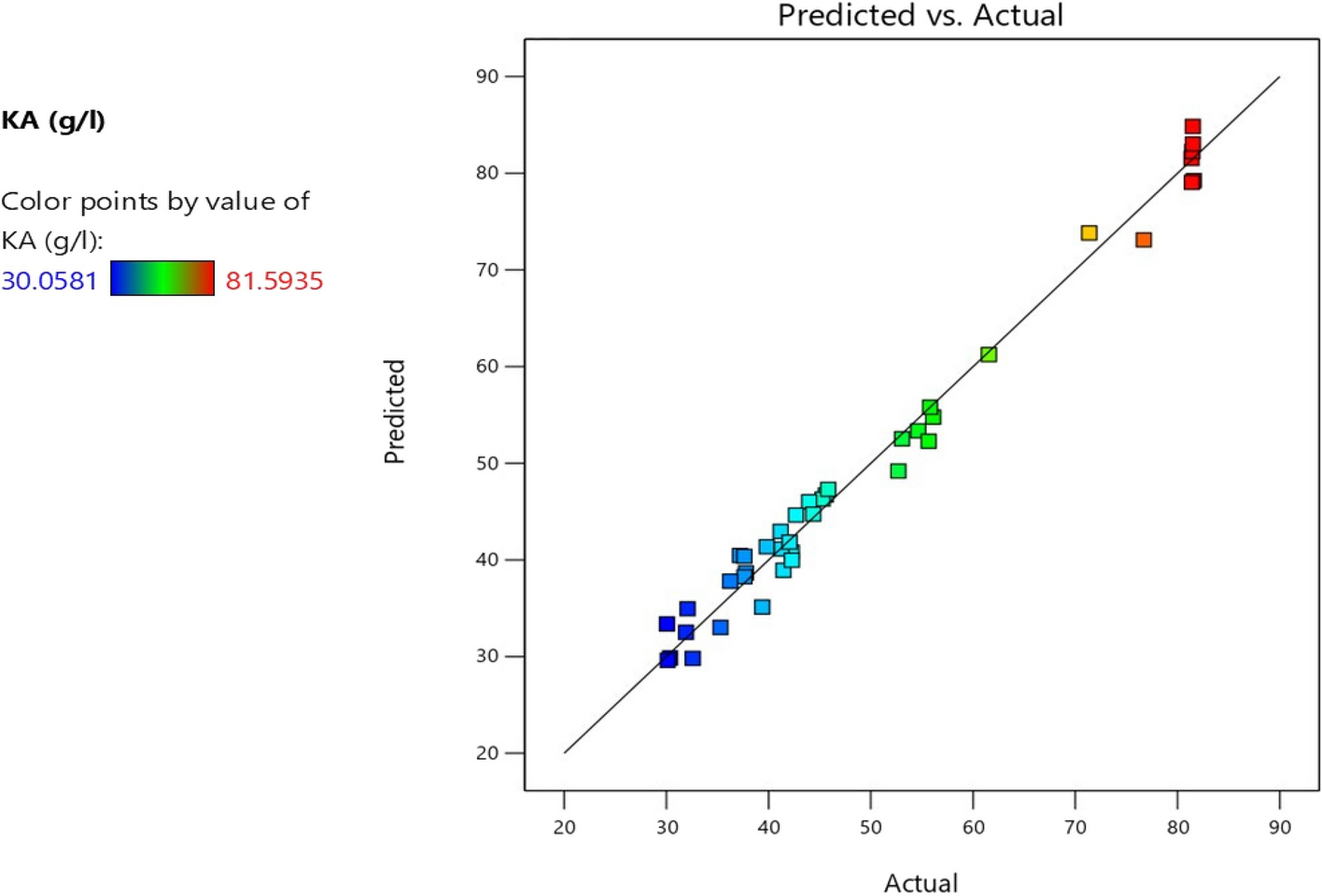
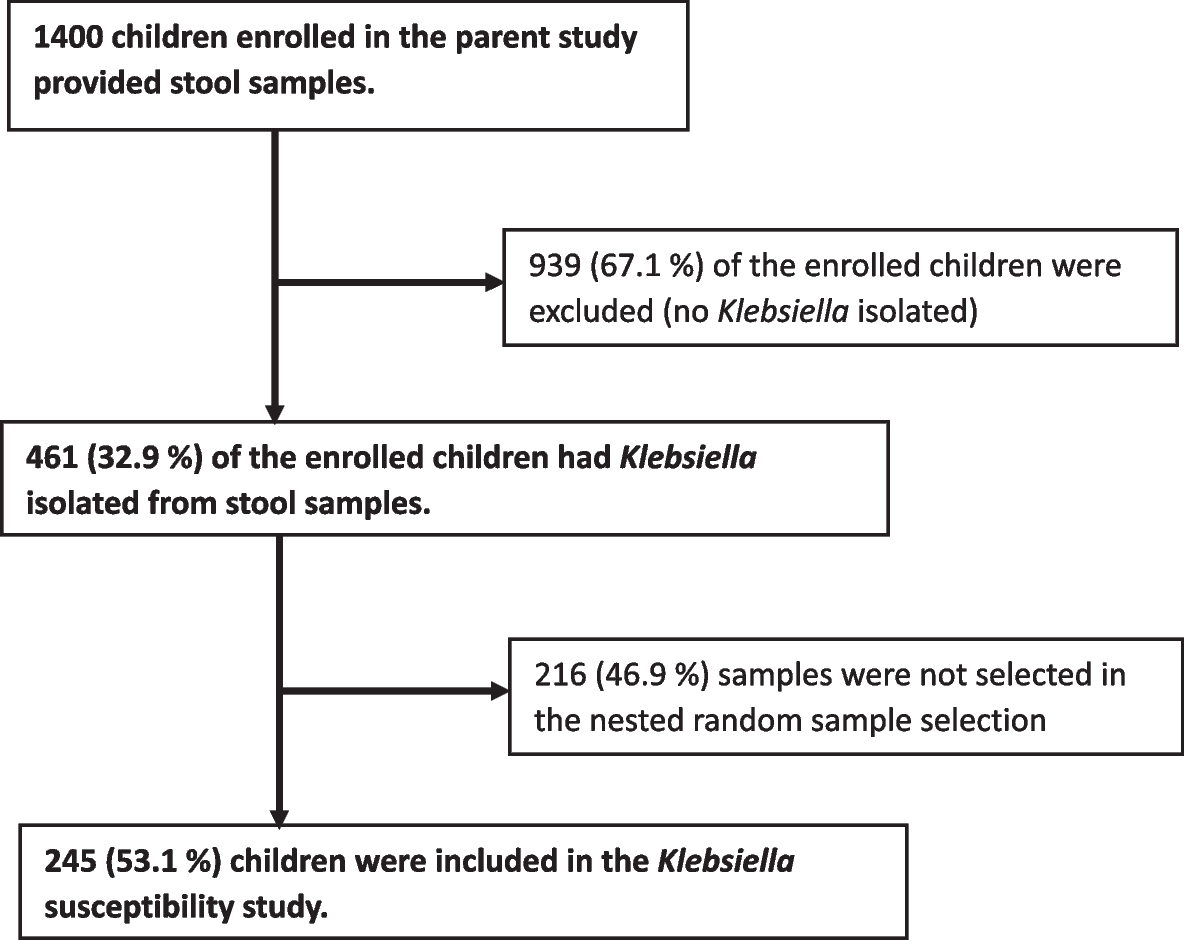
記住我
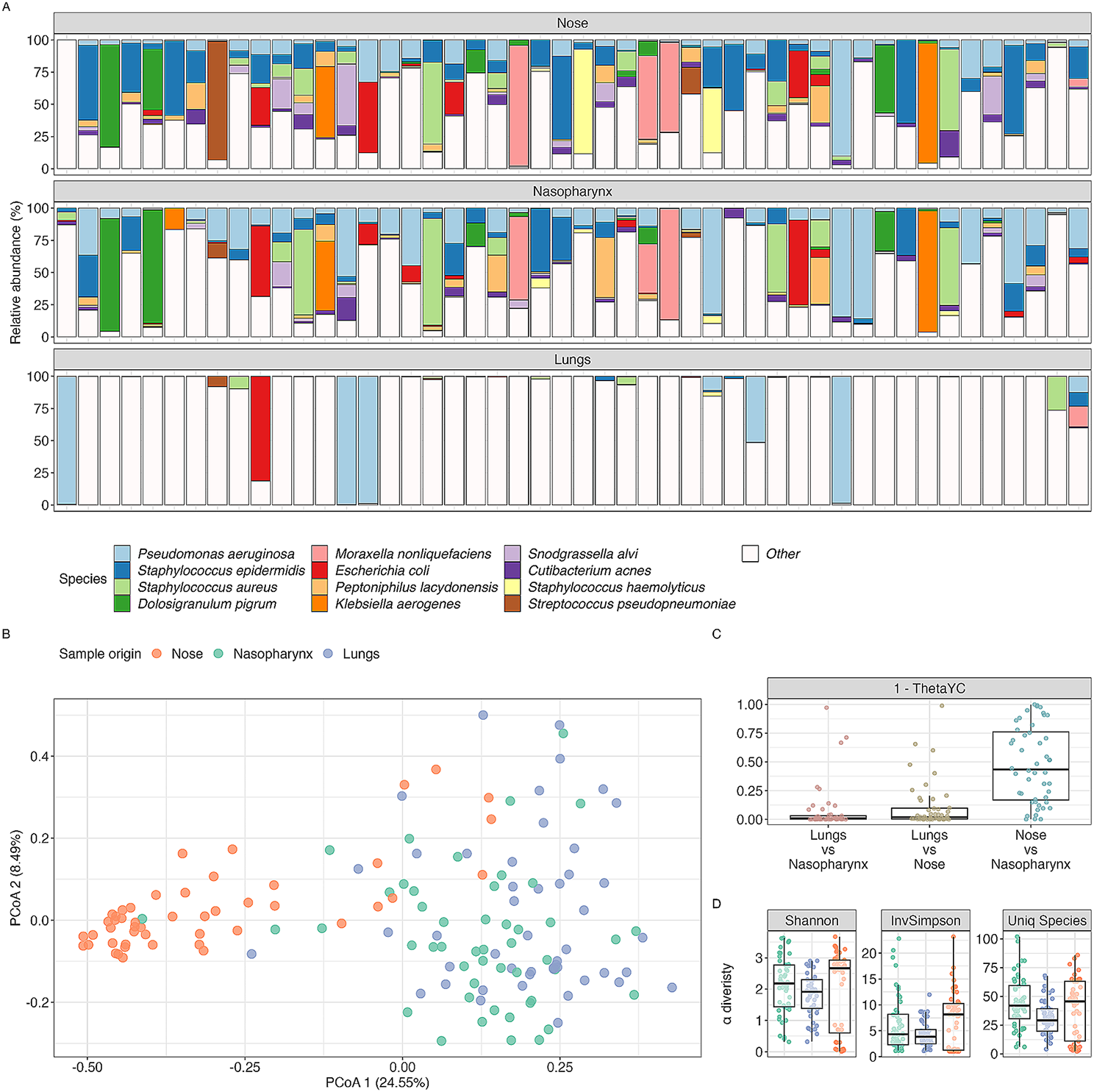

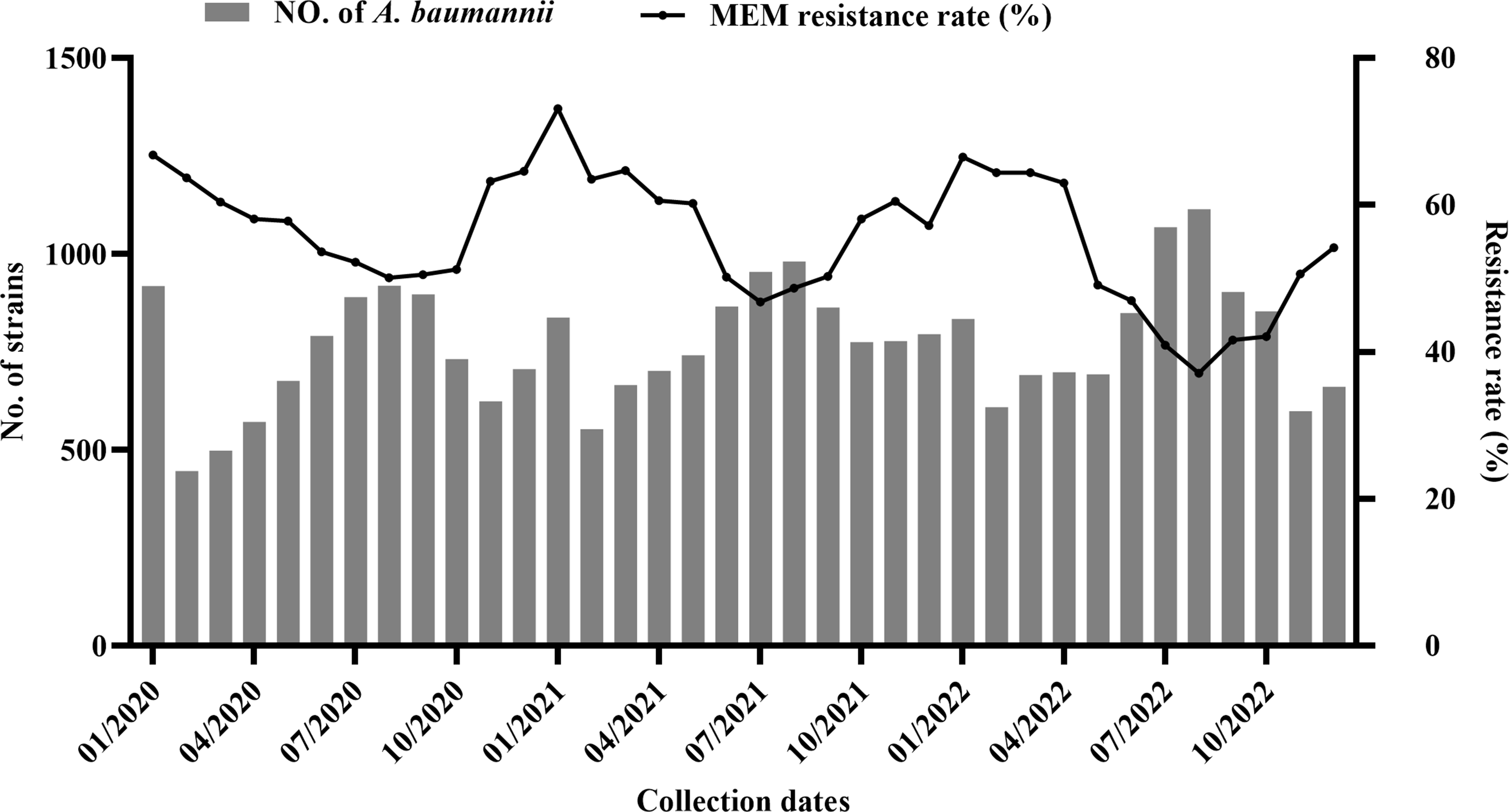
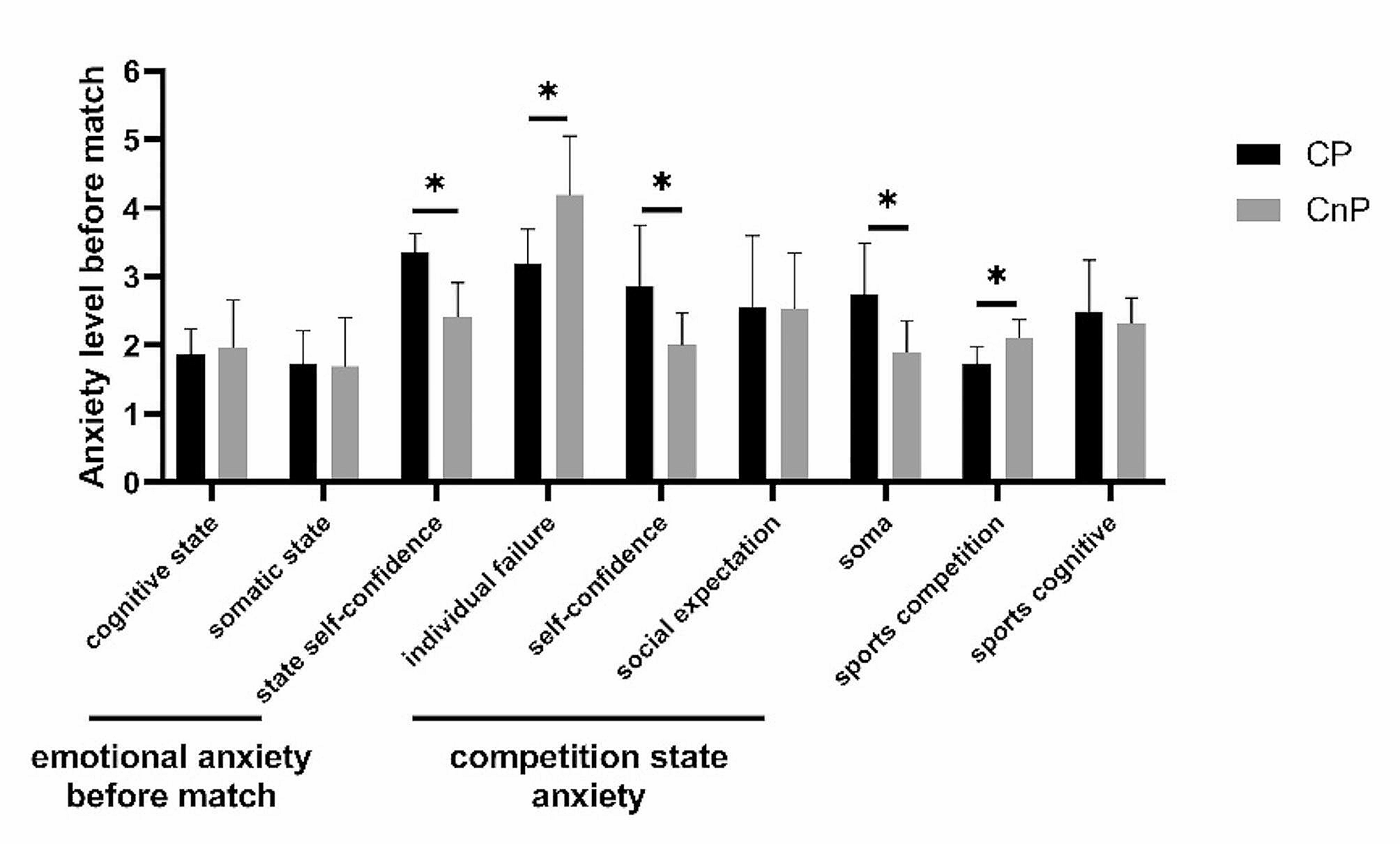
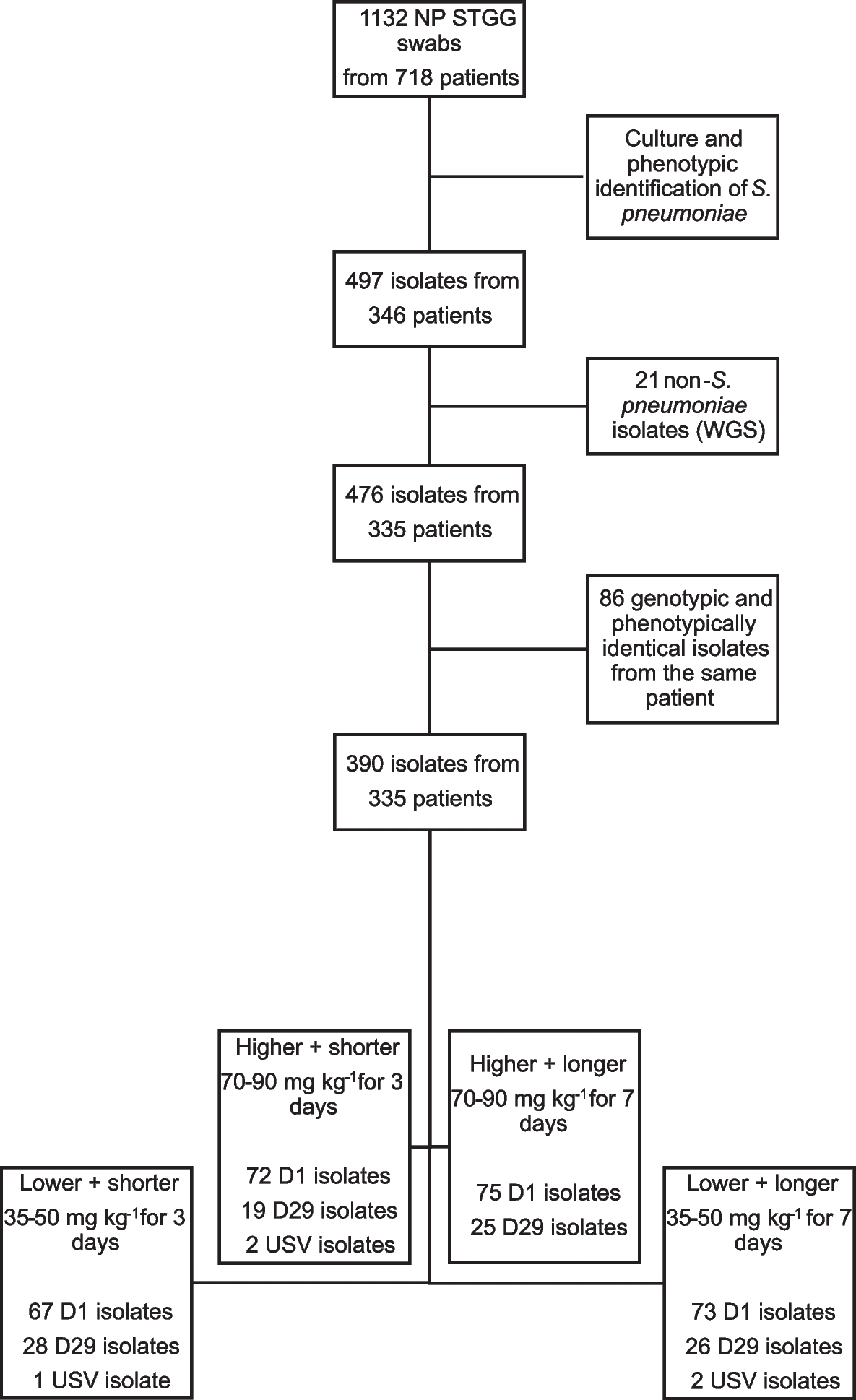


PCR amplification of enterobactin-binding, cell membrane efflux proteins, DNA gyrase, and MurA genes was carried out using two oligonucleotide primers for each gene (Table 1) and the amplicons were examined using agarose gel electrophoresis (Fig. 1A, B). The genes for the enterobactin-binding proteins FepA, FepB, FepC, FepD, and FepG had lengths of 366, 503, 451, 481, and 649 bp, respectively. The purified PCR products of the efflux protein genes AcrB and TolC were 358 and 388 bp, respectively. Additionally, the purified PCR products of the DNA gyrase and MurA genes were 500 and 710 bp in size, respectively.
Table 1 Designed primers and PCR products size of enterobactin binding, efflux proteins, DNA gyrase, and MurA genesFig. 1
Agarose gel electrophoresis of E. coli OQ866153 strain. A, enterobactin-binding and efflux proteins genes amplified by PCR. Lane M: 100 bp molecular weight DNA marker. lanes 1, fepA; 2, fepB; 3, fepC; 4, fepD; 5, fepG; 7, AcrB; and 8, TolC, respectively. B, DNA gyrase and MurA PCR amplicons. Lanes 1, DNA gyrase and 2, MurA
Trojan-Horse strategyPreparation of enterobactin-antibiotic conjugatesThe primary absorbance bands in the FTIR spectrum of pure Ciprofloxacin were observed at 3529, 3375, 3085, 2918, 2687, 2619, 1702, 1622, 1492, 1447, 1383, 1341, and 1269 cm−1 (Fig. S2A). The most significant band, corresponding to the carbonyl group, was found at 1702 cm−1. The IR spectrum bands (941, 1049, 1115, 1279, 1428, 1461, 2941, 3326, and 3560 cm−1) obtained were nearly identical to those of commercially available Fosfomycin sodium salt (Fig. S2B). The characteristic peaks around 1049 and 1115 cm − 1 indicated stretching vibrations of (PO3)2− and (C–O–C), respectively. The IR spectrum of the Fe+3-enterobactin complex resembled that of enterobactin.
Figure 2A shows the presence of the antisymmetric (-NH) stretching band at 3000–3500 cm−1 in enterobactin, both with and without Ciprofloxacin. When Ciprofloxacin was loaded into enterobactin, the disappearance of this band indicated an interaction between the two at the (-NH) position. Figure 2B shows that the antisymmetric stretching bands (-OH) at 1185 and 1500 cm−1 also disappeared in the enterobactin sample, suggesting interaction between enterobactin and Fosfomycin at the (-OH) position. Additionally, complexes prepared at pH 9 were found to be more stable compared to those prepared at pH 4 and 7.
Fig. 2
FTIR spectrum at a 400–4000 cm-1 wavelength range with a resolution of 4 cm−1 showing interaction between enterobactin (Ent) and, (A), Ciprofloxacin (CIP) forming Ent-CPF conjugate (Cnj-1) and (B), Fosfomycin (Fos) forming Ent-Fos conjugate (Cnj-2), at (-NH) and (-OH) positions of enterobactin, respectively
In Vitro antibacterial studiesDetermination of minimum inhibitory concentrations (MICs) by broth microdilution assayTo assess the effectiveness of Fe+3-enterobactin-Ciprofloxacin/Fosfomycin conjugates in inhibiting the growth of E. coli OQ866153, The MIC of each conjugate was examined at various concentrations (1000, 500, 250, 125, 62, 31.25, 7.8, 3.9, 1.9, and 0.97 µg/ml) after a 24-h incubation period. The MIC values for Fe+3-Cnj-1 and Fe+3-Cnj-2 conjugates were 31.25 and 500 µg/ml, respectively. These results indicate that the conjugates have strong activity against E. coli OQ866153. Hence, a study was conducted to investigate the antibacterial mechanism employed by these conjugates against a resistant strain of E. coli (OQ866153).
Time kill assayFigure 3A, B illustrate the time-kill kinetics of Ciprofloxacin and Fosfomycin, both individually and in combination with Fe+3-enterobactin, against E. coli OQ866153. The kinetics of the antibiotics were found to be comparable to those of the bacterial control. Interestingly, the bactericidal activity of Ciprofloxacin was significantly increased at 24 h after treatment with the addition of Fe+3-enterobactin. Initially, there was only a weak inhibition, but this was improved to a bacteriostatic effect. The reductions in bacterial colony-forming units (cfu/ml) at different time points were as follows: 0 h—0.9 log10 cfu/ml, 2 h—2.3 log10 cfu/ml, 4 h—2.54 log10 cfu/ml, 6 h—2.51 log10 cfu/ml, and 8 h—bactericidal effects. This bactericidal effect continued for 12 h and lasted up to 24 h, resulting in a reduction of 3.66 log10 cfu/ml (Fig. 3A). Additionally, the Fe+3-Cnj-2 conjugate demonstrated a bacteriostatic effect for up to 4 h after treatment, resulting in a reduction of 1.3 and 2.7 log10 cfu/ml after 2 and 4 h of incubation, respectively. This was followed by an increase in bactericidal activity, with a reduction of 4 to 5 and 5.7 log10 cfu/ml after 6, 8, and 24 h of treatment (Fig. 3B). It is worth noting that there is no prior research available on the bactericidal effects of these conjugates against antibiotic-resistant E. coli strains. Data analysis was conducted using Graph Pad Prism (Graph Pad Software Inc., San Diego, CA, USA) Version 9.0 for Windows, with one-way ANOVA followed by Dunnett's post hoc test. The results are presented in Tables S1 and S2.
Fig. 3
Time kill kinetics of (A), Fe+3-enterobactin-Ciprofloxacin conjugate (Fe+3-Cnj-1) at 62 µg/ml and (B), Fe+3-enterobactin-Fosfomycin conjugate (Fe.+3-Cnj-2) at 1 mg/ml, compared to their free antibiotics Ciprofloxacin (CPF) and Fosfomycin (Fos), against the E. coli OQ866153
A postantibiotic effect (PAE) assayTable 4 shows the postantibiotic effects (PAE) of Fe+3-Cnj-1, Fe+3-Cnj-2, CPF, and Fos on E. coli, measured as the duration of growth suppression after antibiotic removal. Fe+3-Cnj-1 has a PAE of 3 h, indicating that bacterial growth is inhibited for an additional 3 h after antibiotic exposure. Fe+3-Cnj-2 has a slightly shorter PAE of 2 h compared to Fe+3-Cnj-1. CPF and Fe+3-Cnj-2 both have a PAE of 2 h, suggesting sustained growth inhibition even after antibiotic removal. On the other hand, Fos has a PAE of 0 h, indicating that bacterial growth resumes immediately after the antibiotic is removed (Fig. 4).
Fig. 4
The postantibiotic effect (PAE), or persistent suppression of bacterial growth after brief exposure of a bacterial culture to an antimicrobial agent
DNA-gyrase inhibition assayTo assess the ability of Fe+3-Cnj-1 to inhibit DNA gyrase, a crucial in vitro assay was conducted. This assay is crucial for understanding the antimicrobial efficacy of Ciprofloxacin and its conjugates, as it evaluates their ability to impede the drug target DNA gyrase. The conjugate was tested at different concentrations (0, 5, 15, 25, and 35 μg/ml) and it was found that at 5 μg/ml, there was no suppression of DNA gyrase activity, as evidenced by the presence of supercoiled DNA plasmids. The conjugate showed inhibition of DNA gyrase at concentrations of 15 μg/ml and higher, eliminating supercoiled DNA plasmids at 25 μg/ml. These results demonstrate that Fe+3-Cnj-1 has a higher gyrase inhibitory activity compared to the original Ciprofloxacin drug (35 μg/ml), as shown in Fig. 5.
Fig. 5
Agarose gel electrophoresis for DNA gyrase assay of (A), Fe+3-enterobactin-ciprofloxacin (Fe+3-Cnj-1) conjugate and (B), Ciprofloxacin. + ve, positive control with DNA gyrase and no conjugate; -ve; negative control without DNA gyrase and conjugate
Antibiofilm quantitative assayDifferent concentrations (0, 0.062, 0.125, 0.25, 0.5, 1, and 2 mg/ml) of Fe+3-enterobactin, Fosfomycin, and Fe+3-Cnj-2 were used to determine the level of biofilm formation by E. coli OQ866153. In Fig. 6, it is evident that there was a significant reduction (p < 0.0001) in biofilm formation as the concentration of the conjugate increased. At the MIC level of 0.5 mg/ml, there was a noticeable decrease in biofilm formation. The inhibition percentages of biofilm formation ranged from 4.7% to 100% when the conjugate was added at concentrations of 0.062 to 2 mg/ml. In comparison, Fosfomycin achieved a maximum inhibition percentage of 21.58% at a concentration of 2 mg/ml. The results revealed that the presence of Fe+3-enterobactin enhanced the effectiveness of Fosfomycin by facilitating its cellular uptake via enterobactin-binding proteins (FepABCDG) on the cell membrane, thus overcoming obstacles such as efflux proteins. However, it was observed that Fe+3-enterobactin alone significantly (p < 0.0001) increased biofilm formation, with a percentage of 106.52% at a concentration of 2 mg/ml compared to the untreated control group.
Fig. 6
Effect of Fe+3-enterobactin, Fosfomycin, and Fe+3-enterobactin-Fosfomycin (Fe+3-Cnj-2) on E. coli OQ866153 biofilm formation
Environmental Scanning Electron Microscope (ESEM) analysisThe ESEM analysis demonstrated that Fe+3-Cnj-2 had an inhibitory effect on biofilm formation by E. coli OQ866153. In Fig. 7A, it can be observed that the untreated sample of E. coli OQ866153 attached to the glass surface and formed a biofilm with a dense extracellular polysaccharide (EPS) matrix containing bacterial cells. However, in Fig. 7B, the treated sample with Fe+3-enterobactin exhibited a higher density of an EPS matrix, which wrapped a larger number of proliferated bacterial cells compared to the untreated sample.
Fig. 7
ESEM micrographs of E. coli OQ866153, (A, 20000x), untreated showing EPS of the biofilm structure (orange arrow); (B, 20000x) treated with Fe+3-enterobactin showing increased EPS matrix (orange arrow) and proliferation of cells (blue arrow), and (C; 10000x), treated with Fe+3-enterobactin-Fosfomycin (Fe.+3-Cnj-2) conjugate showing few and disrupted cells (green arrow)
In contrast, when the concentration of the Fe+3-Cnj-2 conjugate was 2 × MIC (1 mg/ml), only a small number of cells were scattered on the glass surface, the biofilm structure had a significantly reduced amount of extracellular polymeric substance (EPS), and the integrity of some bacterial cells was compromised, as illustrated in Fig. 7C. These results suggest that the Fe+3-Cnj-2 conjugate can effectively hinder the formation of biofilms by E. coli OQ866153.
In silico docking st udies Homology ModelThe 3D quaternary structures of FepC, FepD, and FepG were constructed based on their sequence identity with the target proteins. The homology models of FepC, FepD, and FepG were analysed and found to have a 42.19%, 41.06%, and 38.89% identity, respectively, with the iron-hydroxamate import ATP-binding protein FhuC structure of a ferrichrome importer FhuCDB from E. coli (pdb: 7lb8), vitamin B12 import system permease protein BtuC ABC-transporter BtuCD (pdb: 2qi9), and hemin transport system permease protein HmuU of the bacterial heme transporter HmuUV from Yersinia pestis (pdb: 4g1u). The 3D structure of FepC, FepD, and FepG was illustrated in Fig. S3.
Binding Affinity interactionsThe molecular docking study of the enterobactin, Ciprofloxacin, and Fosfomycin ligands to the E. coli cell membrane efflux proteins AcrB (PDB: 1T9U) and TolC (PDB: 1EK9) has been shown in Table 2 and Fig. 8A-F. The docking results indicated that the enterobactin showed the highest binding affinity to protein active sites of AcrB/TolC, with docking scores -6.565/-6.308 kcal/mol, respectively, by forming 8 H-bonds with SER79, THR91, PHE617, GLU683, ASN719, GLU817, and GLU826 of AcrB active site, and 3-H bonds with GLN273 (A), GLN273 (C), and GLY271TolC protein active site. However, both Ciprofloxacin and Fosfomycin showed closely similar binding affinity to the protein active sites of AcrB/TolC with moderate scores of -4.151/-4.113 and -4.964/-4.917 kcal/mol, respectively. These indicated that the cell membrane efflux proteins AcrB and TolC contributed to the export of free iron enterobactin, Ciprofloxacin, and Fosfomycin outside the bacterial cell.
Table 2 The molecular docking study between ligands and the E. coli cell proteinsFig. 8
The 2D and 3D interaction between E. coli proteins and ligands.A, AcrB and Ciprofloxacin; (B), AcrB and Fosfomycin; (C) AcrB and enterobactin; (D), TolC and Ciprofloxacin; (E), TolC and Fosfomycin; (F), TolC and enterobactin; (G), FepA and Fe+3-enterobactin; (H), FepA and Fe+3-enterobactin-Ciprofloxacin; (I), FepA and Fe+3-enterobactin-Fosfomycin; (J), FepB and Fe+3-enterobactin; (K), FepB and Fe+3-enterobactin-Ciprofloxacin; (L), FepB and Fe+3-enterobactin-Fosfomycin; (M), FepC and Fe+3-enterobactin; (N), FepC and Fe+3-enterobactin-Ciprofloxacin; (O), FepC and Fe+3-enterobactin- Fosfomycin; (P), FepD and Fe+3-enterobactin; (Q), FepD and Fe+3-enterobactin-Ciprofloxacin; (R), FepD and Fe+3-enterobactin-Fosfomycin; (S), FepG and Fe+3-enterobactin; (T), FepG and Fe+3-enterobactin-Ciprofloxacin; (U), FepG and Fe+3-enterobactin-Fosfomycin; (V), DNA gyrase and Ciprofloxacin; (W), DNA gyrase and Fe+3-enterobactin-Ciprofloxacin; (X), MurA and Fosfomycin; (Y), MurA and Fe+3-enterobactin- Fosfomycin
Additionally, the Fe+3-Cnj-1/Cnj-2 conjugates exhibited the strongest affinity to enterobactin binding proteins FepABCDG, with respective docking scores of -6.3/-5.605, -6.789/-5.986, -7.120/-7.675, -6.810/-8.016, and -6.360/-6.3136 kcal/mol. Fe+3-Enterobactin followed closely with docking scores of -5.585, -4.388, -7.09, -6.217, and -5.318 kcal/mol, respectively (Table 2 and Fig. 8G-U). Notably, the conjugates of enterobactin with Ciprofloxacin and Fosfomycin exhibited superior binding affinity towards the E. coli DNA supercoiling protein-DNA gyrase and MurA, as indicated by their docking scores (-8.597 and -5.481, respectively), compared to that of the native antibiotics were comparatively lower, with scores of -6.264 and -3.756, respectively (Table 2 and Fig. 8V-Y). These findings suggested that Fe+3-enterobactin could enhance the efficacy of Fosfomycin and Ciprofloxacin by enabling their entry into the cells via FepABCDG proteins on the cell membrane, circumventing the AcrB and TolC membrane barriers.
Assessment of cytotoxic effects on HEK293 human embryonic kidney cells through MTT assayTable 3 shows the IC50 values for Fe+3-Cnj-1, Fe+3-Cnj-2, CPF, and Fos compounds on HEK293 human embryonic kidney cells obtained from the cell viability assay. Fe+3-Cnj-1 had an IC50 of 553.2 μg/ml, while Fe+3-Cnj-2 had an IC50 of 385.7 μg/ml. In comparison, the IC50 values for ciprofloxacin and Fosfomycin were 23.8 μg/ml and 41.4 μg/ml, respectively (Fig. 9).
Table 3 IC50 values of Fe+3-Cnj-1, Fe+3-Cnj-2, CPF, and Fos in HEK293 Human Embryonic Kidney Cells Assessed by MTT AssayFig. 9
Concentration–Response Relationship for Fe+3-Cnj-1, Fe+3-Cnj-2, CPF, and Fos on Cell Viability in HEK293 Human Embryonic Kidney Cells
留言 (0)