The use of push–pull cropping system by small-scale farmers positively impacted soil physicochemical properties, as well as soil and maize-root microbial communities. The positive association of PPT was demonstrated by the enhancement of soil OC, pH, P, N, and B. Additionally, the presence of ecologically important belowground microbial groups involved in soil fertility improvement, decomposition, siderophore production, high carbon sequestration, nutrient cycling, and plant protection in comparison to the Mono cropping system further supported this positive association. These findings can be linked to agroecosystem functions and other ecosystem services, including soil health and maize yield. This section explores the contribution of PPT on belowground ecosystem services provision including soil physicochemical properties, microbiome relative abundance and diversity, shedding light on the system’s role and function in promoting sustainable maize production.
Effect of push–pull cropping system on soil physicochemical properties
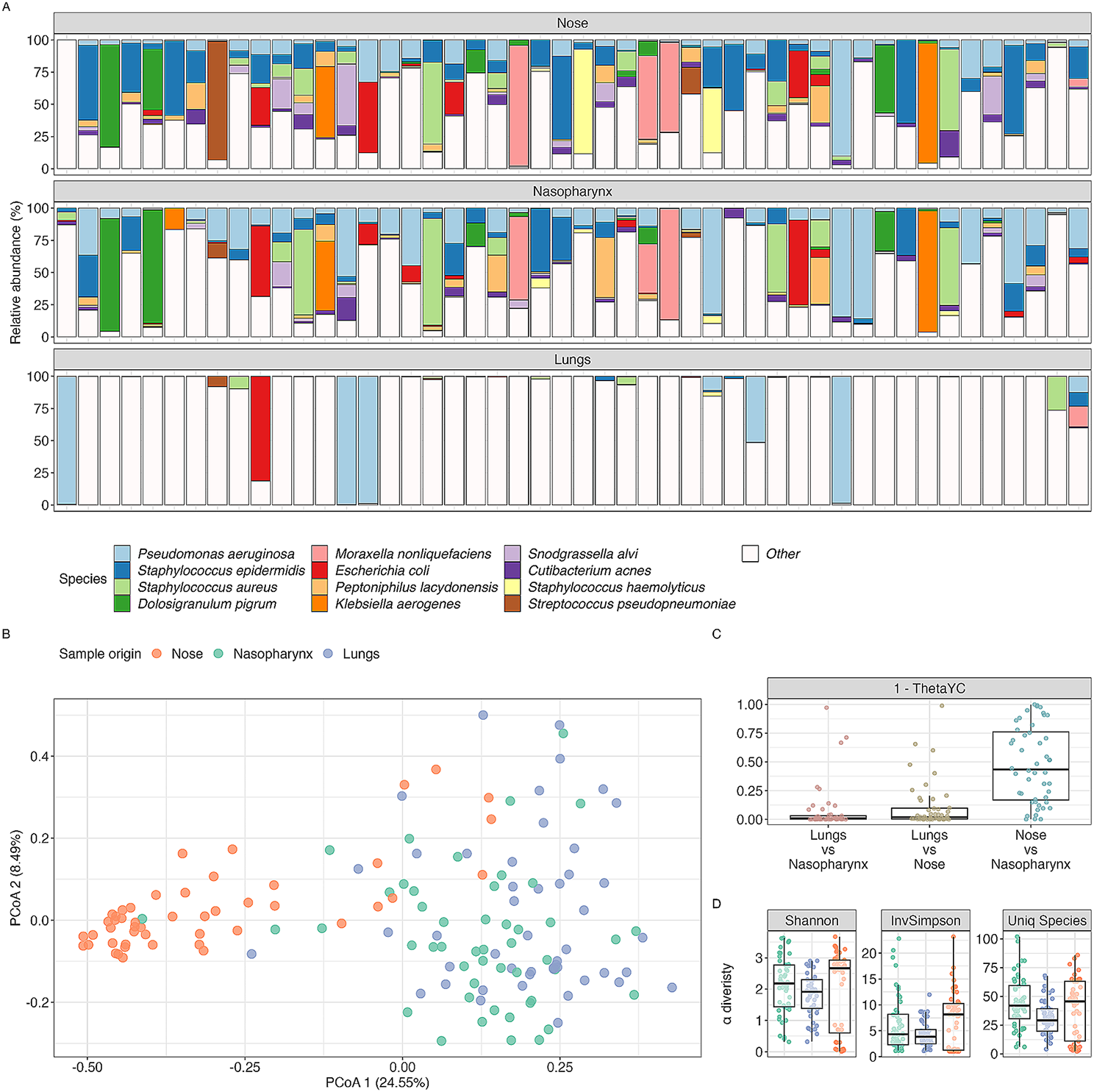
Previously, multiple cropping systems have been shown to influence soil characteristics such as pH, organic carbon, and nitrogen compared to Mono [35, 53]. In this study, we found higher levels of pH, OC, N, and P in soil conditioned by PPT in comparison to the Mono cropping system. A pH below 5.5 can negatively affect plants and pose a significantly threat to the agroecosystem [54]. Given that the pH in the PPT was higher than in Mono, we infer that PPT positively contributes to enhancing soil physicochemical properties and the overall soil health [3, 22]. Frac et al. [55] reported that biotic and abiotic factors, such as soil pH, structure, and nutrient levels, influence the diversity and activity of soil microbes. Our findings regarding the possible influence of cropping systems on both above and belowground abiotic and biotic factors align with the predictions by Drinkwater et al. [3, 5, 54, 56, 57] that crop diversification significantly impacts belowground microbiomes, plant, and soil health, as well as overall productivity.
Impact of push–pull cropping system on soil and maize-root bacterial community
There is growing evidence suggesting that plant diversification impact belowground microbiomes [6, 12, 58]. This study found that PPT cropping systems led to a higher diversity of soil bacterial communities compared to that of the Mono cropping system soil. Notably, the high relative abundances of beneficial bacterial genera such as Sphingomonas, Bacillus, Enterobacter, RB41, Herbaspirillum, Nocardioides, Mitsuaria, Gaiella, Nitrospira, Burkholderia-Caballeronia-Paraburkholderia, Dyella, Enterobacter, and Conexibacter in both the soil and maize-root of PPT systems indicate that PPT favors the proliferation of beneficial bacteria, which improve crop performance and possibly contribute to pest management. Herbaspirillum is a nitrogen-fixing endophytic bacterium that colonizes plant roots and has been shown to positively impact plant growth, and crop yield [59]. Additionally, Bacillus and Enterobacter bacterial genera are potential biofertilizer agents due to their ability to solubilize inorganic phosphate, fix nitrogen, act as biological control agents, carry out bioremediation, and promote plant growth [60, 61]. Sphingomonas, Gaiella, and Dyella play a vital role in promoting plant growth by producing phytohormones and/or inducing changes in phytohormone signalling through volatile organic compound (VOCs), decomposition of lignocellulose, bioremediation of hydrocarbon-contaminated soil, and nutrient cycling in agroecosystem fields [62,63,64]. Additionally, Sphingomonas possesses distinctive capabilities, including the degradation of persistent contaminants, acting as bacterial antagonists to phytopathogenic fungi, and secreting highly beneficial gellan exopolysaccharides [65]. RB41 plays a critical role in regulating the soil carbon cycle and is involved in processing the metabolism of both organic and inorganic nitrogen sources [30, 35, 61]. Furthermore, according to Huang et al. [66], Burkholderia and Mitsuaria genera have a beneficial impact on drought resistance in plants. These bacteria accomplish this by reducing the levels of ethylene, a plant hormone, and producing 1-aminocyclopropane-1-carboxylic acid. Brewer et al. [67] stated that Candidatus Udaeobacter contributes to global hydrogen cycling by utilizing H2. Lazcano et al. [68] found that Nocardioides spp. can act as biocontrol agents for bacterial leaf spots and promote plant growth.
Soil and maize-root from PPT had a greater relative abundance of Streptomyces and Stenotrophomonas, which possess broad biotechnological potential, such as the ability to promote plant growth, production of bioactive secondary metabolites, VOCs, and are promising candidates for biocontrol of phytopathogenic microbes [69]. These characteristics may be attributed to their multiplication rate, ability to produce antibiotics and siderophores, controlled gene expression quorum detection, and synthesis of lipase, chitinase, cellulases, phytohormones, β-1,3-glucanase, and amino acids [70]. Streptomyces spp. can colonize plant root surfaces, survive in various soil types, and produce spores that allow them to persist in extreme conditions. Stenotrophomonas is a potential biocontrol agent against Ralstonia [71, 72]. The presence of these bacterial genera in PPT soils and maize-roots implies that PPT positively influences belowground microbial populations compared to that of Mono. Similar findings have been observed in various other cropping systems, including long-term intercropping systems, push–pull experimental plots, multiple cropping systems, crop rotation, and cover cropping [7, 8, 31, 58]. Nitrospira, which are capable of carrying out nitrification through oxidation of ammonia involving a single organism as opposed to other nitrifying bacteria, which require two different organisms to complete the process [3, 7, 73] were enriched in PPT. These findings imply that PPT influences maize-root microbial populations compared to Mono-root and affects maize-root and soil microbial communities. We also found that the presence of companion crops in a push–pull cropping system had a greater impact on PPT maize-root microbiota such as Streptomyces, Herbasoirillum, Stenotrophomonas, Sphingomonas, Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium, and Dyella compared to Mono-root cropping system. The presence of these beneficial bacteria in the push–pull maize-root may positively contribute to an increase in nitrogen nutrients, carbon sequestration, and biocontrol agent against plant pathogens. This, in turn, can results in improved plant growth due to plant growth-promoting rhizobacterial (PGPR) and siderophores availability, which facilitates iron content in soil and plants from the PPT field. This may lead to higher crop yields in PPT fields compared to that of Mono fields. To better understand the role of different bacterial and fungal species, including those within the same genus, in this cropping system, it is necessary to perform species-level characterization. The finding that Bryobacter, a disease-causing bacterial genus, was more abundant in Mono than in the PPT cropping system shows that Mono cropping systems potentially predispose crops to disease-causing agents.
The high relative abundance of beneficial bacterial species, including Rhizobium phaseoli, Bacillus flexus, Bradyrhizobium elkanii, Paraburkholderia vietnamiensis, Dyella marensis, Enterobacter hormaechei, Herbaspirillum seropedicae, Pseudomonas nitroreducens, Ralstonia pickettii, Sphingomonas paucimobilis, Stenotrophomonas maltophilia, and Variovorax paradoxus, in both the soil and maize-roots within the push–pull cropping systems indicates that this system promotes the proliferation of bacteria that enhance crop performance, improve soil health, water purification, and plant growth, and potentially contribute to insect-pests and disease management. Interestingly, Bacillus flexus possesses the ability to solubilize tricalcium phosphate and hydroxyapatite, making it valuable for biodegradation processes [74, 75]. Bradyrhizobium elkanii produces rhizobitoxine, which acts as a defense mechanism against stress-induced ethylene and plays a significant role in nitrogen fixation [76]. Variovorax paradoxus and Pseudomonas aeruginosa can degrade and/or metabolize N-acyl-homoserine lactones (AHLs) as a carbon source [77]. Chen et al. [78] demonstrated the importance of the complete ethylene signal transduction pathway in enhancing Arabidopsis thaliana growth through the PGPR, Variovorax paradoxus, underscoring the significance of ethylene signalling PGPR activity. Stenotrophomonas maltophilia contributes to bioremediation and nitrogen fixation processes. Interestingly, it contributes to the sulfur cycle and promotes plant growth and health in ecosystems [72, 79]. Sphingomonas paucimobilis enhances antioxidant activity, promotes plant growth, and exhibits biodegradation capabilities [80, 81]. Ralstonia pickettii demonstrates biodegradative abilities through siderophore production, while Pseudomonas nitroreducens produce biosurfactants and solubilizes phosphate [82,83,84]. Herbaspirillum seropedicae, an endophytic diazotrophic PGPR, colonizes various crops (rice, maize, sorghum, and sugarcane) and exhibits beneficial traits such as solubilization of minerals, production of phytohormones, and atmospheric nitrogen fixation [85, 86]. Enterobacter hormaechei has been identified as a potassium solubilizing microbe, showing potential for plant growth and controlling harmful algal blooms [87,88,89,90]. Dyella marensis produces biosurfactants and siderophores, while Paraburkholderia vietnamiensis and Rhizobium phaseoli have shown promise as nitrogen-fixing fertilizers for plant growth [91,92,93].
Impact of push–pull cropping system on soil and maize-root mycobiome
Push–pull cropping system decreased the number of harmful fungal genera. Contrarily, it increased the presence and relative abundance of beneficial belowground fungal genera, such as Mortieralla, Exophiala, Paraboeremia, Bionectria, Clitopilus, Marasmius, Pyrenochaetopsis, and Trichoderma compared to the Mono cropping system. These findings align with previous studies which have demonstrated crop diversification enhance beneficial fungi with a positive impact on agroecosystem productivity [3, 6, 94, 95]. For example, Mortierella spp. has been shown to solubilize phosphate, improve nutrient uptake, and influence soil microbiota, synthesize phytohormones that support plant growth and defense mechanisms [74]. Enriched in PPT, Mortierella and Pyrenochaetopsis spp. are important indicators of soil-root microbiome continuum, enhancing crop yield, disease resistance, and salinity tolerance in tomatoes [90]. Exophiala spp. which was enriched in PPT, has been observed to produce phytohormones and enzymes, promoting plant shoot growth under drought and salinity conditions [15]. Paraboeremia spp. has been demonstrated to increase plant biomass and glycyrrhizin content in Liquorice plants [96], and it can parasitize eggs of the rice root-knot nematode, Meloidogyne graminicola, in in-vitro assays [27, 97]. Bionectria spp. has been shown to decompose plant debris, improve soil health, and act as biological control agents against insect-pests [98]. The volatile antimicrobial compounds produced by this fungus suppress plant pathogens and could be used as an effective biofumigant [99, 100]. Clitopilus spp. produces pleuromutilin, a biologically active compound with potent antimicrobial activity and the ability to increase plant growth through facilitative potassium uptake [101, 102]. Trichoderma spp. found enriched in PPT, is associated with colonizing the rhizoplane, rhizosphere, and plant roots, and produces metabolites with antimicrobial (volatile and non-volatile compounds, cellulose/lignin/cell wall degrading enzymes and antibiotics) and biostimulating properties (phytohormones and phytoregulators) [98, 103]. This fungus has direct and indirect biocontrol potential against soil phytopathogens, increases nutrient solubility, and contributes to plant protection, crop yield, and biofertilization production [104, 105]. Fungal spp. belonging to Ramicandelaber and Robillarda have been reported as decomposers, with Robillarda producing β-1,3/1,4-glucans that contribute to disease resistance in plants [106, 107]. While harmful fungal genera such as Aspergillus, Gibberalla, Neocosmopora, and Curvularia were found to be more enriched in the Mono cropping system compared to that of PPT, it is important to note that not all species within these genera are harmful. Some species within these genera also exist as endophytes. However, some produce toxins; for example, Zearalenone, an estrogenic mycotoxin that is produced by Gibberella spp. causes Gibberella ear rot (GER) in crops like maize, oats, wheat, sorghum, rice, and barley [33, 94, 108]. Neocosmospora, identified as a phytopathogen causing stem rot, adversely affects potato growth and yields, leading to economic losses due to stunted growth, leaf yellowing, and grayish-black stems [109]. Fungal spp. belonging to Curvularia, poses a threat to cereal crops, causing economically burdensome Curvularia leaf spots in maize [110, 111]. Similarly, mycotoxin producing species like Aspergillus, infect various fruits, cereal, and vegetable plants, causing several disorders, reducing seed germination, and impairing root and shoot elongation [32, 112].
Diversity of soil and maize-root microbiome in push–pull and maize-monoculture cropping systems
While annual legume intercropping may temporarily affect belowground microbiome profiles, the impact of perennial companion intercrop, such as Desmodium spp. is expected to be stronger and more resilient, contributing to increased soil and maize-root microbial diversity [3,




留言 (0)