
記住我


The in vitro reconstitution system was prepared by co-culturing bone marrow macrophages from Ctsk-Cre × ROSA26-tdTomato mice with bone nodules formed by osteoblasts from EGFP mice. The isolated osteoblasts from EGFP mice were confirmed to be alkaline phosphatase (ALP)-positive (Suppl. Fig. S1a), and the isolated bone marrow cells from Ctsk-Cre × ROSA26-tdTomato mice were confirmed to differentiate into tartrate-resistant acid phosphatase (TRAP)-positive osteoclasts (Suppl. Fig. S1b).
We first optimized the concentrations for E-64 and ZOL. At a 10 µM or more concentration of E-64, resorption pits decreased as described [9] (Suppl. Fig. S2a, b). The cytotoxicity of E-64 was confirmed through the loss of cellular autofluorescence tdTomato signal due to cell death, and found no significant difference (Suppl. Fig. S2c). From these results, we determined to use 10 µM E-64, as previously described [9]. At ZOL concentrations above 1 µM, almost no osteoclast-like TRAP-positive cells were observed (Suppl. Fig. S2d, e), as reported [22].
To compare the effects of E-64 and ZOL on osteoclast differentiation, we treated osteoclasts with 10 µM E-64 or 1 µM ZOL (Suppl. Fig. S3a). There was no significant difference in the number of TRAP-positive cells with actin rings between the control and E-64 groups, whereas the ZOL-treated osteoclasts showed a significant decrease compared to cells of other groups (Suppl. Fig. S3b). The number of nuclei per osteoclast decreased significantly in both the E-64 and ZOL groups (Suppl. Fig. S3c, d).
To evaluate the effects of E-64 and ZOL on the nodule formation, 10 µM E-64 or 1 µM ZOL was added to the osteoblast differentiation culture, and alizarin red staining was performed after 28 days (Suppl. Fig. S3e). As a result, the ZOL group showed a significant decrease (Suppl. Fig. S3f).
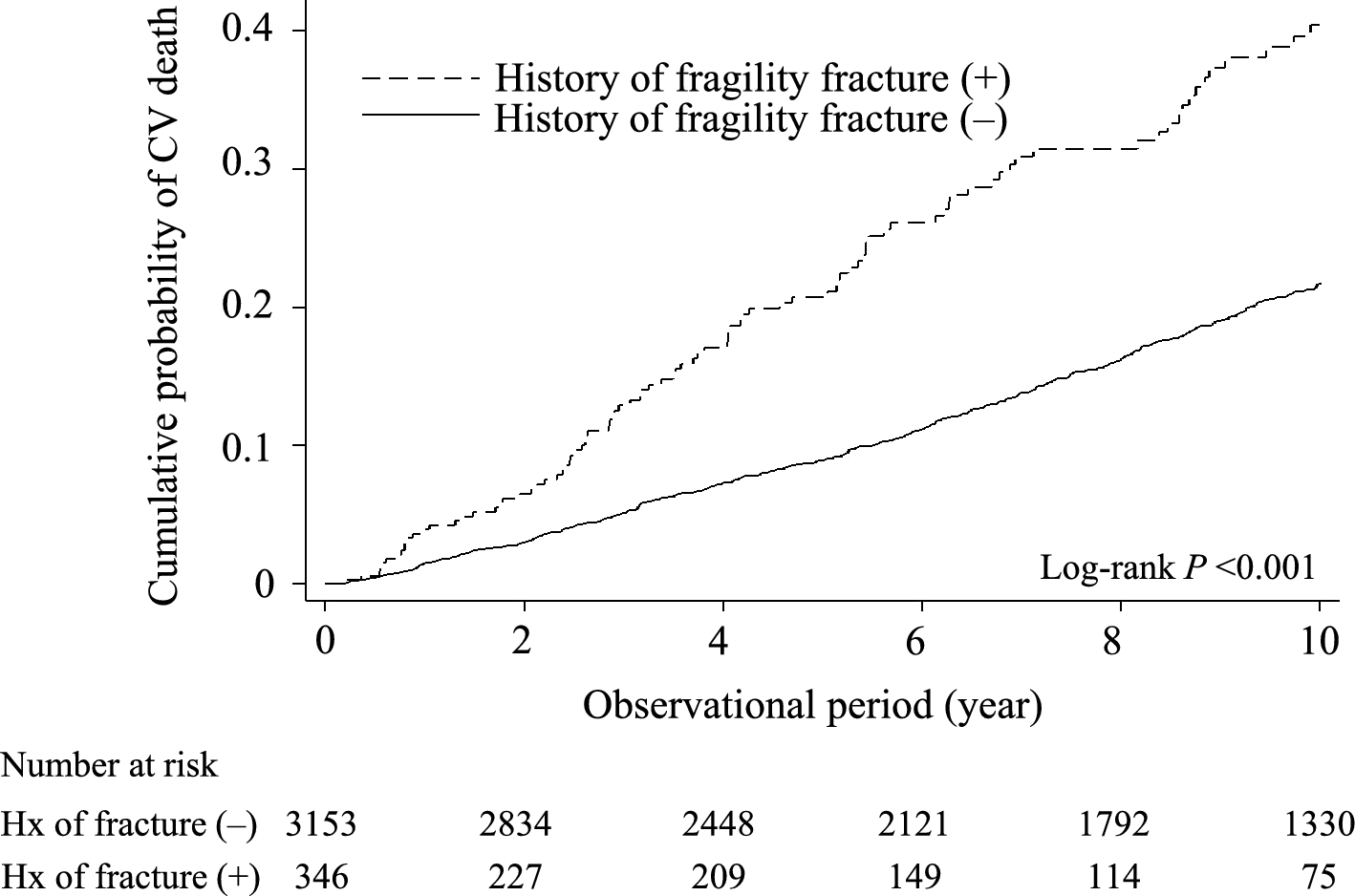
To evaluate the effects of E-64 and ZOL on bone remodeling, we added them to the in vitro reconstitution system. The observations were started at the end of the differentiation culture of osteoblasts (hereinafter referred to as R0). After the first observation, bone marrow macrophages from Ctsk-Cre × ROSA26-tdTomato mice were co-cultured with differentiated osteoblasts (Fig. 1a). The bone marrow macrophages turn red when cathepsin K was expressed during osteoclastogenesis. SHG, which can detect collagen without fluorescence labeling, was used to detect bone nodules formed by osteoblasts.
Fig. 1

The in vitro reconstitution system of bone remodeling. a Schema of the experimental design. Weeks 0, 1, 2, 3, 4, and 5 are referred to as R0, R1, R2, F1, F2, and F3 hereafter. b, c Volumetric views of 3D data acquired by 2-photon microscopy. Maximal impression images shown by IMARIS are presented. b Volumetric views of all channels. Gray: second harmonic generation (SHG; collagen). Green: EGFP (osteoblasts). Red: tdTomato (osteoclasts). c Volumetric views of the SHG channel. To make the images more visible, the brightness of each image was modified using look-up tables (LUTs) and the same values. Scale bar: 100 µm. Yellow arrowheads in the control group: representative resorption pits that were filled with the new matrix created. Dotted yellow circles in E-64: sites at which resorption occurred in the 1st week of the resorption phase and formation occurred in the 2nd week. Dashed magenta circles in E-64: sites at which formation occurred in the 1st week of the formation period, resorption occurred in the 2nd week, and formation occurred again in the 3rd week. d Zoom-up images of osteoclasts shown by NIS-elements AR software (Nikon). e, f Orthogonal views of osteoclasts at R2. The nuclei were stained with Hoechst 33342. The images are via NIS-elements AR. Scale bar: 25 µm. e Volumetric views of all channels. Gray: SHG. Green: EGFP (osteoblasts). Red: tdTomato (osteoclasts). f Volumetric views of the SHG channel. g Quantification of the number of nuclei per osteoclast in panels e and f. Data were analyzed by the Kruskal–Wallis test followed by the Steel–Dwass test. Six fields of each dish were observed, and the experiment was carried out three times (n = 18). Data are mean ± SE. h A scatterplot between the number of nuclei per osteoclast and the osteoclast volume (R2). Spearman’s rank correlation coefficients (R) and p values. n = 18
Resorption pits were formed in the nodules of the control group during the resorption phase (week 0 [R0] to week 2 [R2]) and were later filled with freshly generated matrix in the following formation phase (week 3 [F1] to week 5 [F3], (Fig. 1b, c, yellow arrowheads). The resorption pits in the E-64 group were not as prominent as those of the control group (Fig. 1c). In addition, the synchronicity of the resorption-formation cycles was disturbed by E-64. For example, in some areas, matrix was resorbed at R1 and formed at R2 (Fig. 1c, yellow circles). In other regions (Fig. 1c, magenta circles), the formation occurred at F1, followed by resorption and formation at F2 and F3, respectively. Some resorption pits were not filled (Fig. 1c, red circles). In the ZOL group, changes in the matrix were not as obvious as in the other groups (Fig. 1b, c).
In the control group, multinucleated large osteoclasts appeared at resorption pits during the resorption period, and the matrix was resorbed in a large area (Fig. 1d–f). In the E-64 group, some osteoclasts were detached from the matrix as mentioned in an earlier study [23] (Fig. 1d). Osteoclasts with few nuclei formed small resorption pits (Fig. 1e, f). The number of nuclei was significantly decreased by adding E-64 or ZOL to the co-culture of osteoclasts and osteoblasts, as was the case in the monoculture of osteoclasts (Fig. 1e–g, Suppl. Fig. S3c, d). The number of nuclei per osteoclast and the osteoclast volume (R2) showed a tendency of moderate correlation in the control group (Fig. 1h).
We also confirmed the deposition of ZOL on calcified nodules and its uptake by osteoclasts using fluorescence-labeled ZOL in the in vitro reconstitution system (Suppl. Fig. S3g–n).
The changes in the overall matrix, osteoclasts, and osteoblasts volumeSurface rendering was performed for SHG-, EGFP-, and tdTomato-positive regions for the measurement of the temporal changes in matrix, osteoblasts, and osteoclasts, respectively. The volume and surface area were then calculated. We used the projected area of the SHG at R0 to modify the volume of the SHG, and differences between the values at each time point and those at R0 were calculated (Fig. 2a). First, the temporal changes in the SHG volume were evaluated in each group. The SHG volumes at R2 in the control group were significantly lower than those at R0, R1, F2, and F3, indicating the significant reduction and formation of matrix during the resorption and formation phases, respectively. The SHG volumes at R1, R2, F1, and F2 of the E-64 group were significantly lower than those at R0, in a pattern different from that of the control group. In the ZOL group, the SHG volumes at R1 were significantly lower than at R0 (Fig. 2b).
Fig. 2


Analyses of SHG, tdTomato, and EGFP volumes. a Surface rendering of the SHG positive area (upper panel). The value of the area of maximum SHG projection at week 0 (lower panel) was used to adjust the SHG volumes. Temporal changes in the SHG (b), tdTomato (d), and EGFP (f) volume for each group. Data are mean ± SE. The Friedman test followed by Bonferroni adjustment was used to determine the significance of differences between the values at each time point. c, e, g Comparisons of the three groups’ values at each time point. Data are mean ± SE and were analyzed by the Kruskal–Wallis test followed by the Steel–Dwass test. Control: n = 15, E-64: n = 16, ZOL: n = 17. c SHG, e tdTomato, g EGFP
The differences in the SHG volumes among the three groups at the same time points were also analyzed. The SHG volume in the ZOL group was significantly larger than those of other groups at R1, R2, F1, and F2. There was no significant difference between each group at F3 (Fig. 2c).
We also analyzed the changes in the overall osteoclasts (Fig. 2d, e) and osteoblasts volume (Fig. 2f, g). The osteoclast volume in the E-64 group was significantly larger than those of other groups at F1, and those in the ZOL group were lower than those in the other groups at R1, R2, F2, and F3 (Fig. 2e). The EGFP volume in the ZOL group was significantly lower than those of the control groups at R2 (Fig. 2g).
The regional changes in the matrix volumeOne field of view was divided into 16 ROIs for the analysis of regional changes in the parameters for matrices, osteoclasts, and osteoblasts, and correlations among them [5] (Fig. 3a). To investigate the quantitative and topological correlations between bone resorption and formation, we analyzed the amount of change in the SHG volume during the resorption phase and formation phase in each ROI (Fig. 3b). Bone resorption and bone formation moderately correlated in the control group, indicating that the resorption and formation of the matrix were balanced in each region. In the ZOL group, the corresponding correlations between values were weak. The correlations were negligible in the E-64 group, suggesting that the topological relationship between resorption and formation was disturbed by the inhibition of cysteine proteinases (Fig. 3c, d).
Fig. 3
Analysis of regional changes in the SHG volume. a Sixteen regions of interest (ROIs) in one field of view. b Explanatory diagrams for the values of bone resorption and bone formation, and the scatterplots. Changes in the values during the resorption phase (week 0 [R0] to week 2 [R2]) and the formation phase (week 2 [R2] to week 5 [F3]) are plotted. c, d Spearman’s rank correlation coefficients (R) and p values (c) and scatterplots (d) for bone resorption and bone formation. Control: n = 240, E-64: n = 256, ZOL: n = 272
The correlations between matrix changes and osteoclast volumesTo investigate the effect of osteoclasts on the matrix, we calculated the osteoclast volumes in the ROIs and their correlations with bone resorption/formation. The osteoclast volume peaked at R2 in the control and E-64 groups, and negligible change was observed in the ZOL group, indicating the suppression of osteoclastogenesis (Fig. S4a).
The analysis of the correlations revealed that the osteoclast volume at R2 and the cumulative osteoclast volume (R1 + R2) were strongly correlated with bone resorption in the control group. The correlations were reduced to moderate ones in E-64 group, and almost disappeared in ZOL group (Fig. 4a, b). In the control group, there was a strong correlation between bone resorption and average volume of osteoclasts at R1 and R2 (Fig. 4c).
Fig. 4
Analysis of the correlations between the matrix changes and the osteoclast volume. a Correlations between the changes in the matrix volume and the changes in the osteoclast volume. Spearman’s rank correlation coefficients (R) and p values. b Scatterplots between bone resorption and the cumulative osteoclast volume (R1 + R2). Control: n = 240, E-64: n = 256, ZOL: n = 272. c A scatterplot between bone resorption and the total volume of osteoclasts with a volume over 2000 µm2 at R1 and R2, divided by the number of ROI. Spearman’s rank correlation coefficients (R) and p values. Control: n = 240
The correlations between matrix changes and osteoblast volumesThe osteoblast volumes in each ROI were calculated and their correlations with bone resorption/formation were examined. The osteoblast volume was lowest at R1 in the control and E-64 groups, whereas in the ZOL group, it was lowest at R2 (Suppl. Fig. S5a).
When the correlations between bone formation and osteoblast volume were analyzed, the osteoblast volumes at F1 and F2 showed a strong correlation with bone formation in the control group. On the other hand, only a moderate correlation was observed between bone formation and OB volume at F1 in the ZOL group. Furthermore, in the E-64 group, only the osteoblast volume at F1 was weakly correlated with bone formation (Fig. 5a–c).
Fig. 5
Analysis of the correlations between matrix changes and the EGFP volume. a Correlations between bone resorption/formation and the osteoblast volume. Spearman’s rank correlation coefficients (R) and p values are presented. b, c Scatterplots between bone formation and the osteoblast volume at F1 (b), and F2 (c). Control: n = 240, E-64: n = 256, ZOL: n = 272
The correlations between matrix changes and osteoblast sphericityCuboidal osteoblasts with high sphericity are suggested to be active osteogenic cells [15, 17]. We evaluated the correlations between bone resorption/formation and the sphericity of osteoblasts. From the surface rendering of the SHG and EGFP with heatmaps (Fig. 6a) and the changes in the osteoblast sphericity of each ROI over time (Suppl. Fig. S6a), we did not observe distinct patterns. The analysis revealed that the sphericity of osteoblasts at R2 and bone formation were moderately correlated, suggesting the osteoblasts with relatively high sphericity that existed at the end of the resorption phase affected the subsequent bone formation (Fig. 6b, c).
Fig. 6
Analysis of the correlations between bone resorption/formation and osteoblast sphericity. a Surface rendering of SHG and EGFP shown by the Imaris software. The EGFP-positive area is shown with a heatmap indicating the extent of sphericity. Scale bar: 100 µm. b Correlations between bone formation/resorption and osteoblast sphericity. Spearman’s rank correlation coefficients (R) and p values are presented. c Scatterplots between bone formation and the osteoblast sphericity at R2. Control: n = 240, E-64: n = 256, ZOL: n = 272
The correlations between the osteoblast volume and osteoclast volumeFinally, we analyzed the correlation between osteoblasts and osteoclasts (Fig. 7a, b). In the control group, the osteoclast volume at R2 was moderately correlated with the osteoblast volumes at R2, F1, F2 and F3, and the cumulative osteoclast volume (R1 + R2) also showed a moderate correlation with the osteoblast volume at R2, suggesting the existence of interactions between these two lineages of cells. However, only weak or negligible correlations were observed in the E-64 and ZOL group (Fig. 7a, b).
Fig. 7

Analysis of the correlations between the osteoblast volume and osteoclast volume. a Correlations between the osteoblast volume and osteoclast volume. Spearman’s rank correlation coefficients (R) and p values are presented. b Scatterplots between the osteoblast volume at R2. Control: n = 240, E-64: n = 256, ZOL: n = 272. c–g Orthogonal views of resorption pits in the control and E-64 groups. c Orthogonal views of the control (see Fig. 1b). The images were revealed by NIS-elements AR software. d Cropped pictures of panel c are indicated by red boxes. Yellow arrowheads: spherical osteoblasts. *Flat osteoblasts. e Orthogonal views of E-64 (see Fig. 1b). The images are by NIS-elements AR. f Cropped pictures of panel e are indicated by red boxes. *Flat osteoblasts. To make images more visible, the brightness of each image was modified using LUTs and the same values. Scale bar: 100 µm. g Cropped pictures of panels d and f, respectively, as indicated by blue boxes. Yellow arrowheads: spherical osteoblasts. *Flat osteoblasts
Temporal changes in resorption pitsRepresentative findings that confirm the analysis results with images are provided as Fig. 7c–g. In the control group, osteoclasts expressing tdTomato appeared during the resorption phase and around resorbed matrix. Osteoblasts with high sphericity were observed below the canopy-like structure composed of flat osteoblasts and around the osteoclasts in the resorption pit. In the subsequent formation phase, the resorption pits were then filled specifically. In the adjacent bone surface covered with flat osteoblasts, no resorption or formation was observed (Fig. 7d, g). In the E-64 group, red osteoclasts appeared during the resorption phase, and a shallow resorption pit was formed. The resorption site was covered with flattened osteoblasts. Migrations of spherical osteoblasts were not observed. In addition, the resorption site was not filled by the newly synthesized matrix (Fig. 7f, g).
留言 (0)