
記住我
Subarachnoid hemorrhage (SAH), which is primarily induced by ruptured intracranial aneurysms, remains a remarkable clinical challenge with prominent incidence and death rates worldwide [1]. Previous studies have mainly focused on delayed cerebral ischemia or cerebral vasospasm following SAH [2,3]. Nonetheless, in recent years, research has demonstrated that the commonest risk factor for disability and death in aneurysmal SAH patients is early brain injury (EBI) [4,5]. The sharply increased intracranial pressure after hemorrhage causes global ischemia and hypoxia, which eventually leads to brain tissue damage and neurological dysfunction [6,7]. Although considerable efforts have shed light on the mechanisms involved in pathological events associated with SAH, the potential pathophysiological mechanisms are still poorly understood. Recently, several reports have found that stimulator of interferon genes (STING) levels are significantly elevated and significantly exacerbate neuroinflammatory events in different central nervous system disorders [8,9].
STING, also known as mediator of interferon regulatory factor 3 activation (MITA), methionine–proline–tyrosine–serine (MPYS), endoplasmic reticulum (ER) IFN stimulator (ERIS) and transmembrane protein 173 (TMEM173), contains 379 amino acids, consists of five transmembrane patterns and is mainly located in the ER, where it contributes to the production of type I IFN. Meanwhile, the STING has the essential roles in innate immune response against inflammation [9]. As we all know, one of the fundamental reactions of the innate immune responses is the release of proinflammatory cytokines, including IL-1β, processed by the NLRP3 inflammasome [10].
Alpha-lipoic acid (ALA), a dithiol antioxidant, is an important cofactor in pyruvate dehydrogenase and α-ketoglutarate dehydrogenase in the mitochondria [11]. The metabolic antioxidant lipoic acid is a low molecular weight substance that is absorbed from the diet and crosses the blood–brain barrier. Although the metabolic effects of ALA have been known for more than 50 years, its exogenous effects have only recently become known. Several studies have demonstrated that ALA supplementation reduces oxidative stress and restores reduced levels of other antioxidants in the body [12]. ALA affects many cellular processes and modulates the impairment of a variety of CNS diseases, including Parkinson, and Alzheimer diseases, traumatic brain injury (TBI), cerebral ischemia, seizures and autoimmune encephalomyelitis and SAH [11,13].
In the light of these findings, the purpose of this study was to define whether ALA alleviates SAH-induced EBI via the inhibition of STING activation and subsequent NLRP3 inflammasome reduction.
Materials and methods AnimalsSeventy-two adult Sprague–Dawley rats (male, 250–280 g) purchased from Bestest (Zhuhai, China) were used in our study. All study protocols were authorized by the Institutional Animal Care and Use Committee of Guangdong Medical Hospital (Zhanjiang, China). All animal management was conducted in accordance with the National Institutes of Health published Guide for the Care and Use of Laboratory Animals (7th Edition). Rats were housed in cages with ad libitum feeding at controlled room temperature (22 ± 2 °C) and humidity (60–80%) under a 12 h/12 h light/dark cycle.
SAH modelThe establishment of the SAH model was operated based on previous studies with some modification [14]. In brief, rats were anesthetized with 40 mg/kg 1% pentobarbital sodium via intraperitoneal injection. Under a microscope, a 3–0 monofilament was inserted via the left external carotid artery into the internal carotid artery and perforated the bifurcation of the anterior and middle cerebral arteries. Sham-operated rats underwent the same procedures without perforation. Temperature of the rats was maintained at 37 ± 0.5 °C during the surgery. At last, rats were monitored for recovery and then returned to cages.
Animal grouping and drug administrationALA (Sigma, T5625-5G, purity >95%) was dissolved in normal saline and injected in a volume of 30 mg/kg through caudal vein into rats 30 min after SAH operation. SR-717 (MedChemExpress, New Jersey, USA, HY-131454), an agonist of STING, was injected in a volume of 30 mg/kg intraperitoneally into rats 30 min after SAH operation.
The rats were divided into four groups randomly: (1) the sham group, rats underwent sham operation; (2) the SAH group, rats underwent SAH; (3) the SAH + ALA group, rats underwent SAH and injection of ALA solution; (4) the SAH + ALA + SR-717 group, rats underwent SAH, injection of ALA and SR-717. All the rats were euthanized by cervical dislocation after anesthesia after SAH operation for 24 h, and brain was isolated for further exploration after behavioral tests.
Neurological scoringAfter SAH for 24 h, the neurological score was measured using the beam balance and modified Garcia’s methods. Beam balance score was conducted to assess athletic ability [15]. The score of the modified Garcia test has six tests that evaluate spontaneous activity, symmetry in the movement of four limbs, climbing ability, response to vibrissae stimulation, forepaw outstretching and body proprioception. The neurological core ranges from 3 to 18.
Hematoxylin–eosin staining and Nissl stainingRats were transcardially perfused with 500 mL 4% paraformaldehyde at 24 h after SAH, as formerly reported [16]. After fixation in 4% paraformaldehyde, dehydration and paraffin-embedding, 4-μm-thick consecutive coronary sections of rat brain were prepared. After dewaxing, the sections were stained with hematoxylin–eosin (H&E) solution according to a commercial kit (C0105S, Beyotime Biotechnology, Shanghai, China). Nissl’s staining was conducted based on the previous study [17]. In short, the slices were incubated in the toluidine blue solution (G1036, Servicebio, Wuhan, China) for 8 min. Images were obtained via a light microscope (Leica, DM2500, Vizsla, Germany).
Immunohistochemistry stainingAt 24 h post-SAH, rats were anesthetized and transcranial perfused with 250 mL PBS (0.1 M, pH 7.4) followed by 500 mL 4% paraformaldehyde. Then, 4- μm-thick coronal brain sections (n = 4/group) were deparaffinized in xylene, rehydrated via an alcohol gradient and washed with PBS (0.01 M, pH 7.4). Immunohistochemical staining was performed based on the commercial kit (SPN-9002, ZSGB-Bio, Beijing, China). Antigen retrieval was performed with citrate-EDTA antigen retrieval solution (P0086, Beyotime, China) in a microwave oven lasting for 25 min. After that, endogenous peroxidase activity was blocked employing 0.3% H2O2 for 10 min and then washed with PBS. After blocked by 5% goat serum for 15 min, slices were incubated overnight at 4 °C with the mouse monoclonal anti-STING antibody (1 : 100, NBP2-24683, Novus). Adjacently, sections were incubated with biotinylated goat antimouse IgG secondary antibody for 15 min and later with HRP-streptavidin reagent. Ultimately, immunoreactivity was visualized using 3,3-diaminobenzidine (Boster), followed by restaining with hematoxylin. Images were obtained via a light microscope (Leica, DM2500, Germany).
Western blotWestern blot was performed according to our previously protocols [16]. Briefly, cerebral cortical tissue was harvested from the injured side of SAH, then homogenized and centrifuged. The detergent compatible protein assay kit (Bio-Rad, Hercules, California) was used to determine the protein content. Equal amounts of protein (30 μg) were loaded into the wells of the SDS-PAGE gel. After electrophoresis, the protein was transferred to a nitrocellulose membrane. The membrane was blocked for 2 h using a nonfat dry milk buffer and incubated overnight at 4 °C with the primary antibodies. The following primary antibodies were used: anti-STING (1 : 1000, NBP2-24683, Novus), anti-NLRP3 (1 : 1000, Abcam, ab270449), anti-ASC (1 : 1000, Abcam, ab309497), anti-caspase-1 (1 : 1000, Abcam, ab207802), anti-IL-1β (1 : 1000, Abcam, ab315084), anti-β-actin (1 : 1000, Abcam, ab213262). The membrane was incubated for 1 h with secondary antibodies at room temperature. The membrane was exposed to X-ray film, and band densities were analyzed using Image J software. To facilitate comparisons between groups, band density values were normalized to the mean value for the sham group.
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) stainingTUNEL staining was performed using a TUNEL kit (Roche Inc., Indianapolis, Indiana). Briefly, brain sections were incubated with TUNEL reaction mixture for 1 h at room temperature. After washed with PBS three times, DAPI solution was used for the cell nucleus (stained blue) staining for 5 min at room temperature. At last, a fluorescence microscope was used to observe TUNEL positive cells (Leica, DM2500, Germany).
Data statisticsAll data are presented as the means ± SD. Data analysis was performed using SPSS 23.0 (IBM SPSS, Chicago, Illinois). Statistical significance among three or more groups was analyzed by one-way analysis of variance followed by Tukey’s post hoc test, while that between two groups was calculated using Student’s t test. P < 0.05 was regarded to have statistical significance.
Results ALA attenuated neurobehavioral deficits and reduced histological damage after SAH at 24 hRats in the SAH groups showed severe behavioral deficits (Fig. 1a) and imbalance (Fig. 1b), compared with rats in the Sham group at 24 h post-SAH. Treatment of ALA improved behavioral performance as well as coordination and balance of movement. H&E staining and Nissl staining (Fig. 1c) were also used to evaluate the effect of ALA administration at 24 h post-SAH; the results indicated that ALA administration alleviated histological impairments.
 Fig.1:
Fig.1: The effect of ALA treatment on neurological function and neuronal degeneration after SAH. (a) Representative images of the bottom of rat brains from sham and SAH at 24 h. (b,c) ALA treatment significantly improved neurological function. (d) Representative images of H&E and Nissl stainings showing alterations in brain injury after ALA treatment. Data are expressed as the mean ± SD (n = 4/group). *P < 0.05 vs. sham group; #P < 0.05 vs. SAH group. ALA, alpha-lipoic acid; H&E, hematoxylin–eosin; SAH, subarachnoid hemorrhage.
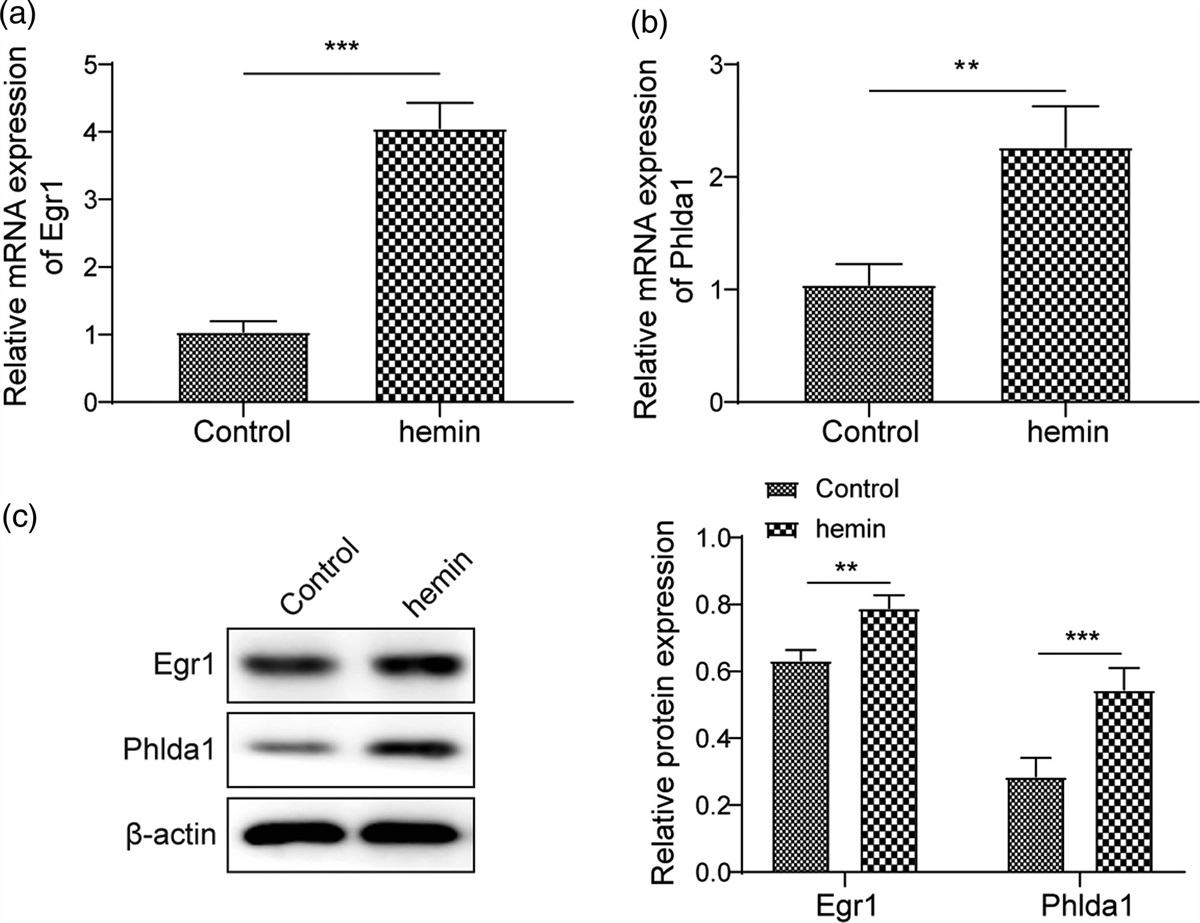
ALA inhibited expression of STING-NLRP3 protein at 24 h post-SAHImmumohistochemical staining images showed that STING expression levels were significantly elevated at 24 h after SAH. STING expression levels of ALA treatment group, however, were declined at 24 h post-SAH when compared with the SAH group (Fig. 2a). In addition, we performed Western blotting to further explore the correlation between STING and inflammation. Western blot results showed that expression levels of STING and NLRP3 protein were significantly increased after SAH, whereas they decreased in the ALA-treated group (Fig. 2b,c).
 Fig. 2:
Fig. 2: Effect of ALA on STING-NLRP3 protein expression at 24 h after SAH. (a) ALA treatment decreased the expression of STING in brain tissue at 24 h post-SAH; (b) Western blot of NLRP3 and STING at 24 h post-SAH in cerebral cortex. (c, d) Data are expressed as the mean ± SD (n = 4/group). *P < 0.05 vs. sham group; #P < 0.05 vs. SAH group. ALA, alpha-lipoic acid; SAH, subarachnoid hemorrhage; STING, stimulator of interferon genes.
STING agonist SR-717 inhibited the neuroprotection and antiapoptotic effects of ALA, aggravating neurological deficits after SAHTo confirm STING inhibition in the neuroprotective effect of ALA, STING agonist SR-717 was used. Modified Garcia Score and Beam Walking Score were performed to evaluate the neurological function in different groups. Compared with ALA treatment group, SR-717 + ALA treatment showed severe behavioral deficits (Fig. 3b) and imbalance (Fig. 3b) close to SAH group. H&E staining and Nissl staining (Fig. 3c) were also used to evaluate the inhibition effect of SR-717 on ALA neuroprotection effect. Meanwhile, the TUNEL staining revealed that apoptotic neurons were notably increased in both SAH and ALA + SR-717 groups (Fig. 4). Taken together, these data indicated that SR-717 reversed the cerebroprotective effect of ALA on EBI following SAH via STING activation.
 Fig. 3:
Fig. 3: Effect of SR-717 on the neuroprotective effect of ALA. (a,b) SR-717 reduces behavioral scores and reverses the neuroprotective effects of ALA on SAH. Data are expressed as the mean ± SD (n = 6/group). *P < 0.05 vs. sham group; #P < 0.05 vs. SAH group. $P < 0.05 vs. SAH + ALA group. (c) Representative images of H&E and Nissl stainings in the Sham, SAH, SAH + ALA, and SAH + ALA + SR-717 groups. ALA, alpha-lipoic acid; HE, hematoxylin–eosin; SAH, subarachnoid hemorrhage.
 Fig. 4:
Fig. 4: SR-717 inhibits the antiapoptosis of ALA. (a) Representative images of TUNEL in the Sham, SAH, SAH + ALA and SAH + ALA + SR-717 groups. (b) Data are expressed as the mean ± SD (n = 4/group). *P < 0.05 vs. sham group; #P < 0.05 vs. SAH group. $P < 0.05 vs. SAH + ALA group. ALA, alpha-lipoic acid; SAH, subarachnoid hemorrhage; TUNEL, terminal deoxynucleotidyl transferase dUTP nick-end labeling.
ALA mediates NLRP3 inflammasome activation via STING signalingWestern blot results showed that coadministration of ALA and SR-717 significantly increased the levels of STING, NLRP3, ASC, cleaved caspase-1 and mature IL-1ß, compared with the SAH + ALA group (Fig. 5).
 Fig. 5:
Fig. 5: SR-717 inhibits regulation of the spine-NLRP3 signaling pathway by ALA. (a) Western blot of NLRP3 and STING at 24 h post-SAH, and relative expressions of ASC, caspase-1 and IL-1β in cerebral cortex. (b–f) Data are expressed as the mean ± SD (n = 4/group). *P < 0.05 vs. SAH + vehicle group. ALA, alpha-lipoic acid; SAH, subarachnoid hemorrhage; STING, stimulator of interferon genes.
DiscussionThe results of the present study demonstrate that ALA significantly reduces cell death in brain tissue and ameliorates neurological deficits after SAH. In addition, our report found that activation of STING and subsequent NLRP3 and related inflammatory factors were involved in the process of EBI after SAH. Moreover, ALA inhibited the STING-NLRP3 pathway activation, thereby attenuating the inflammatory process and ameliorating neurological deficits after SAH, whereas these effects were abolished by STING agonist SR-717. Taken together, our findings suggested that ALA administration, targeting STING-NLRP3 pathway activation, might be an effective therapeutic strategy after SAH.
Inflammation has been widely accepted as a crucial factor in SAH-induced EBI, which is eminently related to poor neurologic function outcome [18–20]. STING, also known as TMEM173, MPYS, MITA or ERIS, is a protein on the ER that can be activated by immune-stimulatory DNA and initiate type I interferon responses and was identified in 2008 [21,22]. Subsequently, several DNA sensors, including interferon-gamma inducible protein 16 (IFI16), were found to support STING activation [23]. Studies have revealed the participation of the STING pathway in unregulated inflammatory processes, TBI, spinal cord injury, hypoxic–ischemic encephalopathy and multiple neurodegenerative disorders [24–26]. Recently, evidence showed that the STING levels are notably increased within 12 h and its activation play a crucial role in the inflammatory mediates by microglia [8]. Hence, therapies targeting STING activation might attenuate EBI and improve neurological function following SAH.
Traditionally, ALA (thioctic acid) is an organosulfur component produced from plants, animals and humans. It has various properties, among them great antioxidant potential, and is widely used as a racemic drug for diabetic polyneuropathy-associated pain and paresthesia [27]. ALA’s pharmacological effects are primarily related with its antioxidant activity, but ALA has also demonstrated interesting cardiovascular, cognitive, antiaging, detoxifying, anti-inflammatory, anticancer and neuroprotective properties [27,28]. Recently, the protective pharmacological effects of ALA have been demonstrated in the treatment of neurological diseases, such as ischemia, seizures, TBI and SAH [13,29–31]. Emerging evidence suggests that ALA has anti-inflammatory properties in neurodegenerative disorder such as Parkinson, Alzheimer Disease and Systemic Lupus Erythematosus [27,32], but few study on SAH. In our study, ALA was found to reduce the expression of STING, NLRP3, ASC, caspase-1 and IL-1β, alleviate the neural necrotic cell death and brain injury and improve the neurological function following SAH. Moreover, coadministration of ALA and STING agonist reversed the neuroprotective effects of ALA only administration after SAH. These data indicated that ALA alleviates EBI after SAH by the inhibition of STING activation and subsequently NLRP3 with the downstream inflammasome activation.
It is important to note that there are several limitations in our present study. First, the antiapoptotic and antioxidant properties of ALA both have been reported; therefore, we cannot exclude the possibility that the property of antioxidant also played a role in the neuroprotective effect of ALA. Second, our study showed that STING activation induced the process of inflammation, but which neural cells were responsible for this process needs to be further explored. In addition, we studied the short-term effects of ALA after SAH, but the side effects of ALA warrant further study.
ConclusionALA inhibits STING activation and subsequent reduction of NLRP3 inflammasomes and attenuates early brain damage after SAH, providing a potential therapeutic intervention for SAH.
AcknowledgementsThis study was supported by Medical Scientific Research Foundation of Guangdong Province of China (No. B2023271), Science and Technology Planning Project of Maoming City (No. 2022153).
Conflicts of interestThere are no conflicts of interest to declare.
References 1. Laiwalla AN, Ooi YC, Liou R, Gonzalez NR. Matched cohort analysis of the effects of limb remote ischemic conditioning in patients with aneurysmal subarachnoid hemorrhage. Transl Stroke Res 2016; 7:42–48. 2. Francoeur CL, Mayer SA. Management of delayed cerebral ischemia after subarachnoid hemorrhage. Crit Care 2016; 20:277. 3. Geraghty JR, Testai FD. Delayed cerebral ischemia after subarachnoid hemorrhage: beyond vasospasm and towards a multifactorial pathophysiology. Curr Atheroscler Rep 2017; 19:50. 4. Al-Mufti F, Amuluru K, Smith B, Damodara N, El-Ghanem M, Singh IP, et al. Emerging markers of early brain injury and delayed cerebral ischemia in aneurysmal subarachnoid hemorrhage. World Neurosurg 2017; 107:148–159. 5. Rass V, Helbok R. Early brain injury after poor-grade subarachnoid hemorrhage. Curr Neurol Neurosci Rep 2019; 19:78. 6. Fujii M, Yan J, Rolland WB, Soejima Y, Caner B, Zhang JH. Early brain injury, an evolving frontier in subarachnoid hemorrhage research. Transl Stroke Res 2013; 4:432–446. 7. Fang YJ, Mei S-H, Lu J-N, Chen Y-K, Chai Z-H, Dong X, et al. New risk score of the early period after spontaneous subarachnoid hemorrhage: for the prediction of delayed cerebral ischemia. CNS Neurosci Ther 2019; 25:1173–1181. 8. Hu X, Zhang H, Zhang Q, Yao X, Ni W, Zhou K. Emerging role of STING signalling in CNS injury: inflammation, autophagy, necroptosis, ferroptosis and pyroptosis. J Neuroinflammation 2022; 19:242. 9. Balasubramaniam A, Srinivasan S. Role of stimulator of interferon genes (STING) in the enteric nervous system in health and disease. Neurogastroenterol Motil 2023; 35:e14603. 10. Liu J, Zhang X, Wang H. The cGAS-STING-mediated NLRP3 inflammasome is involved in the neurotoxicity induced by manganese exposure. Biomed Pharmacother 2022; 154:113680. 11. Ersahin M, Toklu HZ, Cetinel S, Yüksel M, Erzik C, Berkman MZ, et al. Alpha lipoic acid alleviates oxidative stress and preserves blood brain permeability in rats with subarachnoid hemorrhage. Neurochem Res 2010; 35:418–428. 12. Moini H, Packer L, Saris NE. Antioxidant and prooxidant activities of alpha-lipoic acid and dihydrolipoic acid. Toxicol Appl Pharmacol 2002; 182:84–90. 13. Alpha-lipoic acid protects against reperfusion injury following cerebral ischemia in rats. Mol Med Rep 2015; 11:3659–3665. 14. Liu W, Li R, Yin J, Guo S, Chen Y, Fan H, et al. Mesenchymal stem cells alleviate the early brain injury of subarachnoid hemorrhage partly by suppression of Notch1-dependent neuroinflammation: involvement of Botch. J Neuroinflammation 2019; 16:8. 15. Ruan J, Yao Y. Behavioral tests in rodent models of stroke. Brain Hemorrhages 2020; 1:171–184. 16. Fan H, Ding R, Liu W, Zhang X, Li R, Wei B, et al. Heat shock protein 22 modulates NRF1/TFAM-dependent mitochondrial biogenesis and DRP1-sparked mitochondrial apoptosis through AMPK-PGC1alpha signaling pathway to alleviate the early brain injury of subarachnoid hemorrhage in rats. Redox Biol 2021; 40:101856. 17. Hu E, Li T, Li Z, Su H, Yan Q, Wang L, et al. Metabolomics reveals the effects of hydroxysafflor yellow A on neurogenesis and axon regeneration after experimental traumatic brain injury. Pharm Biol 2023; 61:1054–1064. 18. Chamorro A, Meisel A, Planas AM, Urra X, van de Beek D, Veltkamp R. The immunology of acute stroke. Nat Rev Neurol 2012; 8:401–410. 19. Lucke-Wold BP, Logsdon AF, Manoranjan B, Turner RC, McConnell E, Vates GE, et al. Aneurysmal subarachnoid hemorrhage and neuroinflammation: a comprehensive review. Int J Mol Sci 2016; 17:497. 20. Osgood ML. Aneurysmal subarachnoid hemorrhage: review of the pathophysiology and management strategies. Curr Neurol Neurosci Rep 2021; 21:50. 21. Ishikawa H, Barber GN. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008; 455:674–678. 22. Ishikawa H, Ma Z, Barber GN. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009; 461:788–792. 23. Unterholzner L, Keating SE, Baran M, Horan KA, Jensen SB, Sharma S, et al. IFI16 is an innate immune sensor for intracellular DNA. Nat Immunol 2010; 11:997–1004. 24. Abdullah A, Zhang M, Frugier T, Bedoui S, Taylor JM, Crack PJ. STING-mediated type-I interferons contribute to the neuroinflammatory process and detrimental effects following traumatic brain injury. J Neuroinflammation 2018; 15:323. 25. Gamdzyk M, Doycheva DM, Araujo C, Ocak U, Luo Y, Tang J, et al. cGAS/STING pathway activation contributes to delayed neurodegeneration in neonatal hypoxia-ischemia rat model: possible involvement of LINE-1. Mol Neurobiol 2020; 57:2600–2619. 26. Wang YY, Shen D, Zhao L-J, Zeng N, Hu T-H. STING is a critical regulator of spinal cord injury by regulating microglial inflammation via interacting with TBK1 in mice. Biochem Biophys Res Commun 2019; 517:741–748. 27. Salehi B, Selamoglu Z, Mileski SK, Pezzani R, Redaelli M, Cho WC, et al., Insights on the use of alpha-lipoic acid for therapeutic purposes. Biomolecules 2019; 9:773. 28. Biewenga GP, Haenen GR, Bast A. The pharmacology of the antioxidant lipoic acid. Gen Pharmacol 1997; 29:315–331. 29. Freitas RM. The evaluation of effects of lipoic acid on the lipid peroxidation, nitrite formation and antioxidant enzymes in the hippocampus of rats after pilocarpine-induced seizures. Neurosci Lett 2009; 455:140–144. 30. The effects of alpha lipoic acid on cerebral vasospasm following experimental subarachnoid hemorrhage in the rabbit. Turk Neurosurg 2011; 21:527–533. 31. Toklu HZ, Hakan T, Biber N, Solakoğlu S, Oğünç AV, Sener G. The protective effect of alpha lipoic acid against traumatic brain injury in rats. Free Radic Res 2009; 43:658–667. 32. Bharat S, Cochran BC, Hsu M, Liu J, Ames BN, Andersen JK. Pre-treatment with R-lipoic acid alleviates the effects of GSH depletion in PC12 cells: implications for Parkinson’s disease therapy. Neurotoxicology 2002; 23:479–486.
留言 (0)