
記住我
Bordetella pertussis is the bacterial causative agent of pertussis or whooping cough. Despite widespread vaccination programs, B. pertussis is still in circulation with an estimated 24 million cases and 160,000 deaths in children younger than 5 years in 2014 (Yeung et al., 2017). To reduce this public health burden, two major types of vaccine are in use. The first one is a whole-cell pertussis vaccine (wP) composed of inactivated B. pertussis. The second is an acellular pertussis vaccine (aP), composed of purified B. pertussis virulence factors. The most commonly purified antigens found in aP are the pertussis toxin (PT) and the filamentous hemagglutinin (FHA). Some aPs are supplemented with pertactin (PRN) and fimbriae (Fim; Edwards and Decker, 2018). Vaccine developers continuously work on process optimization of wP and aP vaccines, especially antigen production during B. pertussis culture (Dehottay and Goffin, 2013; Dehottay et al., 2015). Despite their critical roles during the production of pertussis vaccines, the biological mechanisms and metabolic pathways, which take place during culture process, are not fully understood.
In 2004, the FDA launched the process analytical technology (PAT) initiative, which aims at improving the robustness and consistency of pharmaceutical manufacturing processes. PAT provides a strategy to obtain maximum control of product and process based upon scientific understanding (Food and Drug Administration, 2004). To this end, with a focus on ensuring the robustness of their production, vaccine manufacturers are constantly facing new challenges. As any other bioproduction processes, industrial antigen production is at risk of being hampered by low yield and batch failure. In addition, vaccine producers face a fluctuating demand for their products and must provide consistent scale-up and scale-down processes. Moreover, health authorities recommend the suppression of raw material from animal and, more recently, non-chemically defined supplements (Food and Drug Administration, 2019). To incrementally adapt to these challenges, access to an extensive knowledge of B. pertussis physiology is a prerequisite, particularly regarding the general metabolism and the molecular regulatory mechanisms associated with vaccine antigen production.
Bordetella pertussis displays several defective metabolic pathways. It is reported to have dysfunctional glycolysis and requires niacin supplementation for growth. Bordetella pertussis only uses cysteine as sulfur source and its tricarboxylic acid (TCA) cycle has long been considered partially non-functional (Jebb and Tomlinson, 1957; Thalen et al., 1999; Parkhill et al., 2003). However, B. pertussis TCA cycle was recently shown to be entirely functional in the Tohama I strain using chemically defined SS-medium (Izac et al., 2015). In addition, some B. pertussis strains are also able to use thiosulfate as a sulfur source. Thus, B. pertussis metabolic capabilities are strain-dependent (Branco dos Santos et al., 2017).
The regulation of B. pertussis virulence during bacterial culture has been well documented over the past decades. Several studies uncovered the regulatory mechanisms of the main virulence regulon BvgASR in specific B. pertussis strains, using virulence modulators such as niacin or MgSO4 in flask culture models followed by transcriptomic analyses (Hot et al., 2003; Moon et al., 2017; Coutte et al., 2020). Recently, two additional two-component systems, RisAK and PlrSR, were shown to be involved in B. pertussis virulence regulation, in vitro and in vivo, respectively (Cróinín et al., 2005; Coutte et al., 2016; Bone et al., 2017). Despite these findings which were mostly obtained using flask culture models, a lot remains to be uncovered to have a global picture of B. pertussis metabolism and to fully understand B. pertussis virulence regulation during culture.
With the development of OMICs technologies, deep molecular longitudinal analyses of culture processes are now accessible, allowing the monitoring of thousands of molecules interacting across different omics. Such multi-omics approaches have been successfully applied in a wide range of biological fields including microbiology (Hasin et al., 2017; O’Donnell et al., 2020). Through a study combining proteomic and microarray analyses on a chemostat model, proteomics was shown to be a valuable method to monitor and assess the quality of B. pertussis antigens along the wP upstream production process (Metz et al., 2017). In a subsequent study, a genome-scale metabolic model combined to an end-point extracellular metabolomic analysis provided a new insight into the metabolic capabilities of B. pertussis and led to a > 2-fold improvements in pertussis toxin production (Branco dos Santos et al., 2017). More recently, the combination of longitudinal and multi-omics analyses demonstrated its value for the characterization and optimization of industrial culture bioprocesses as shown with the fungus Aspergillus niger (Lu et al., 2018) or with the bacterium Zymomonas mobilis (Zhang et al., 2019). Despite initial very promising investigations (Nakamura et al., 2006; Van De Waterbeemd et al., 2009; Branco dos Santos et al., 2017), such an extensive longitudinal multi-omics study has never been reported before on B. pertussis using experimental conditions mimicking industrial culture processes.
Here, we report the first longitudinal investigation of a small-scale B. pertussis bioreactor model combining transcriptomic, proteomic, lipidomic, and metabolomic analyses. Data obtained with this model that intends to mimic industrial process, provide an insight into the metabolism and the regulation networks of B. pertussis during culture in bioreactors.
Materials and methods Bacterial strainA colony of B. pertussis Tohama I (ATCC BAA-589) was isolated from a lyophilized commercial stock solution cultured on a Bordet Gengou agar (BD, #254400) plate. From this colony, a master seed bank was obtained using a 48 h cultivation on Bordet Gengou agar plate and aliquoted in Bovine Serum Albumin—Saccharose Phosphate Glutamate buffer. A working seed bank was next produced from the master seed bank cultured using a chemically defined SS-medium in a fermenter. The culture was then harvested in exponential growth phase and aliquoted following the addition of 20% of glycerol and stored at −80°C.
Bacterial cultureSix batch cultures of Bordetella pertussis (Figure 1), strain Tohama I (ATCC®BAA-589), were performed in a chemically defined medium derived from the Stainer-Scholte (SS) medium as previously described (Izac et al., 2015). This medium reproduces the composition of casein hydrolysate-containing medium used in industrial processes (Imaizumi et al., 1983; Quentin-Millet et al., 1987). Preculture and cultures were carried out in small scale biofermenters (Ambr® 250, Sartorius), in a batch mode at 36°C, pH 7.3, and 26% of dissolved oxygen (DO). Anti-foam C (Dow Corning™) was used to control foaming and pH was regulated by automatic addition of 10% (v/v) H3PO4. Dissolved oxygen was controlled by a regulation cascade of stirring (350–1,300 rpm), air injection (10–110 ml.min−1), and dioxygen injection (0–80 ml.min−1).
Figure 1. Experimental plan of the longitudinal multi-omics study of Bordetella pertussis culture.
Briefly, preculture was inoculated from the frozen working seed bank. After 22 h of growth, an appropriate volume of preculture was centrifuged to replace preculture supernatant with fresh medium. Fermenters were inoculated at 0.1 of optical density at 650 nm wavelength (OD650). Sampling was performed for quality control (QC) testing, antigen quantification, and omics analyses. End of the cultures were determined by a sudden decrease of carbon emission rate (CER), and an increase of dissolved oxygen.
Culture quality controlDuring cultures, temperature, pH, dissolved oxygen, foaming, and CER were monitored using the Ambr® 250 platform. Growth monitoring was ensured by measuring the OD650 (V-630 Spectrophotometer, Jasco®) and flow cytometry viability assays. Numeration and bacterial viability were analyzed using a Guava® easyCyte™ flow cytometer (Luminex) with Live/Dead® BacLight™ kit (Thermo Fisher Scientific). At the end of cultures, samples were plated on tryptic soy agar (TSA; BD, #254086) plate to control the culture purity and the absence of avirulent phenotype.
Vaccine antigens quantificationThe quantification of vaccine antigens was performed at 1 h, 4 h, 8 h, 12 h, 18 h 45 min, 22 h, and 26 h of culture by enzyme-linked immunosorbent assay (ELISA; Figure 1). At the end of the cultures, antigen concentrations were measured in triplicate for each culture. Run acceptance criteria was defined, for all final antigen concentrations, at CV < 15% between bioreactors.
Briefly, 125 μl of crude harvest was lysed by incubation with 50% v/v of lysis buffer (Xtractor™buffer, Clontech) during 20 min at room temperature and stored at −80°C. The lysis solution contained 20 U.ml-1 of Turbo DNase (Thermo Fisher, #AM2239), 0.4 mg.ml−1 of lysozyme (Sigma-Aldrich, #L3790), and ethylenediaminetetraacetic acid (EDTA)-free protease inhibitor cocktail (cOmplete™, Roche). Antigen quantifications were performed by sandwich ELISA on B. pertussis lysed suspension using proprietary monoclonal antibodies against specific proteins: PT, FHA, PRN, and Fimbriae 2 (Fim2). Briefly, 96 well plates were coated with monoclonal antibodies anti-PT, anti-FHA, anti-PRN, or anti-Fim2, at a concentration of 3, 2, 2, and 2.5 μg.ml−1, respectively. After a saturation step, samples were diluted to appropriate concentration. After incubation and wash steps, plates were incubated with biotinylated secondary antibodies anti-PT, anti-FHA, anti-PRN or anti-Fim2. Then, a solution of streptavidine-HRP (Jackson Immunoresearch) was added to the plates. Plates were revealed with tetramethylbenzidine (Tebu-bio) incubation during 20 min at room temperature. The reaction was stopped by the addition of 1 N HCl. Optical density at 450 nm (OD450) and 620 nm (OD620) wavelengths were measured using a microplate reader PHERAstar™ FS (BMG Labtech). OD620 values were subtracted to OD450 to consider plastic absorption. Antigen concentrations were determined using SoftMax® Pro (v6.5) software (Molecular Devices) with a standard curve obtained using purified antigens as reference standards.
Sampling methods for multi-omics analysesA total of six batch cultures were used in this study. Three of the six cultures were used to collect samples after 1 h of culture. Due to the low level of biomass at the beginning of the culture, large volumes of the bacterial cultures had to be collected to allow multi-omics analyses. Thus, to prevent a bias due to a change of the culture conditions in the subsequent time points, those three bioreactors were stopped. The three remaining bioreactors were sampled after 4 h, 8 h, 12 h, 18 h 45 min, 22 h and 26 h of culture for all omics analyses (Figure 1).
For transcriptomic analyses, a volume corresponding to 1 OD650 in 1 ml (1 ODu) was sampled and incubated for 1 h at room temperature with 33% v/v of RNAlater (Invitrogen). Then, samples were centrifuged for 10 min at 4,000 g at 4°C and supernatants were discarded.
For proteomic analyses, 1 ODu was sampled and centrifuged for 10 min at 4,000 g at 4°C. Supernatants were eliminated, and pellets were washed twice with 5 ml of cold PBS.
For lipidomic analyses, 4 ODu were sampled and mixed with 50% (v/v) of quenching solution (40% ethanol/0.8% NaCl at −25°C; Spura et al., 2009).Then, samples were immediately centrifuged for 10 min at 4,000 g and 4°C and supernatants were eliminated.
For extracellular metabolomic analyses, at least 300 μl were sampled, centrifuged for 10 min at 4,000 g at 4°C and supernatants were collected.
For all omics analyses, samples were immediately frozen in dry ice and stored at −80°C until analysis.
Transcriptomic data generation and analysisBacteria pellets were lysed in 200 μl of Tris-EDTA buffer (Invitrogen™) with 15 mg.ml−1 of lysozyme (Thermo Fisher Scientific) and 1 mg.ml−1 of proteinase K (Qiagen) for 10 min at 400 rpm, at room temperature. Then, RNA extraction was performed on column using RNeasy® mini kit (Qiagen) including an on-column DNase digestion. A second step of DNase digestion was performed by DNase TURBO™ (Thermo Fisher Scientific). RNA purity was assessed using NanoDrop™ 2000 (Thermo Scientific™). RNA integrity was analyzed by TapeStation 4200 (Agilent) using the High Sensitivity RNA ScreenTape. Only samples with a RINe over 7 were selected. RNA was quantified using Qubit™ 2.0 Fluorometer (Thermo Fisher Scientific) with the Qubit™ RNA high sensitivity kit (Invitrogen™).
Libraries were prepared in triplicate for each sample with the Sciclone® G3 NGSx iQ™ liquid handling workstation (PerkinElmer) by using the Universal RNA-Seq kit (Tecan Genomics, 2021) customized for B. pertussis. Briefly, cDNA was synthetized by reverse transcription and mechanically fragmented with the Covaris® M220 Focused-ultrasonicator™ for 90 s at 50 W, 10% duty factor, and 200 burst per cycle, to obtain 400 bp (base pair) cDNA fragments. Fragments were, then, purified using Agencourt® beads, followed by end repair, adaptor ligation, and sense strand selection steps. Ribosomal RNAs (rRNA) were depleted using AnyDeplete technology (Tecan Genomics) with specific B. pertussis rRNA probes. Libraries obtained after PCR amplification (18 cycles) were characterized by TapeStation with D5000 ScreenTape (Agilent), normalized at 10 nM, pooled, diluted at 4 nM and, denatured according to Illumina NextSeq500 protocol A (Illumina, 2022). Denatured libraries were diluted at 1.8 pM and 1 μl of PhiX at 20 pM was added as control. Then, three sequencing runs were launched, with 1.3 ml of this mix on a NextSeq® 500 platform (Illumina) using 75-bases single-end protocol on a High Output flow cell (Illumina).
Following base calling steps by sequencer software (Illumina), quality control of the sequencing run was carried out using Sequencing Analysis Viewer (SAV, Illumina, version 2.4.7) regarding reads quality and cluster density. After demultiplexing steps, sequencing QC was performed on raw data with fastqc tool (Babraham Bioinformatics, 2019) and visualized using multiQC tool (Ewels et al., 2016).
Raw data were imported in Array Studio (v11.0) software (OmicSoft Qiagen). Reads were trimmed to remove low quality bases with default parameters and mapped to the NCBI genome of the strain of B. pertussis Tohama I (RefSeq accession number NC_002929.2; Parkhill et al., 2003). Finally, QC was performed on aligned reads. Count table was generated with default parameters. Transcripts were normalized using RLE (implemented in DESeq2 R package; Love et al., 2014), those with low abundance (mean normalized expression under 10) were filtered out.
Proteomic data generation and analysisDue to technical limitations, simultaneous analyses of the secretome and cellular proteome could not be achieved in this study. Proteomic analyses were thus solely performed on cellular proteome. Frozen cell pellets were thawed by the addition of 130 μl lysis buffer [4% SDS, 50 mM Tris–HCl (pH 7.5)]. After resuspension, the bacteria lysates were boiled at 95°C on a thermoshaker for 10 min. Following cooling at room temperature, lysates were transferred to Covaris tubes and subjected to ultrasonication on a Covaris® M220 Focused-ultrasonicator™ for 90 s at 75 W, 20% duty factor, and 200 burst per cycle. Next, samples were transferred to new 1.5 ml Eppendorf tubes and centrifuged for 10 min at 10,000 g at 20°C. Protein amounts were determined using a Pierce™ Rapid Gold BCA Protein Assay Kit (Thermo Fisher Scientific).
Aliquots (50 μg) of each protein suspension were next reduced during 20 min using a 10 mM final concentration of dithiothreitol and alkylated with a 20 mM final concentration of iodoacetamide for 20 min in the dark, both at 37°C. Subsequently, sample buffer was immediately removed using the Single-Pot Solid-Phase-enhanced Sample Preparation (SP3) as described previously (Müller et al., 2020). Briefly, 500 μg of Sera-Mag SpeedBeads Hydrophilic (GE Healthcare) were added to samples (10/1 w/w beads-to-protein ratio), then acetonitrile (ACN) was added to a final concentration of 75%. The beads were agitated with a thermoshaker for 10 min at 500 rpm at room temperature followed by 2 min of incubation in a magnetic rack. The supernatant of each sample was removed, and beads were washed twice with 200 μl of 80% ethanol and once with 200 μl of neat ACN. Beads were resuspended in 90 μl of 100 mM ammonium bicarbonate solution containing 1 μg of Trypsin/Lys-C Protease Mix (1/50 w/w enzyme-to-protein ratio; Thermo Fisher Scientific). Enzymatic digestion was carried out for 2 h on a thermoshaker at 37°C with 500 rpm agitation. Following digestion, the samples were sonicated for 5 min in a sonic bath, centrifuged for 1 min, and incubated for 2 min on the magnetic rack, then 90 μl was transferred to a new plate and 10 μl of 20% ACN / 5% trifluoroacetic acid was added to quench enzymatic digestion.
Reverse phase separation of 1 μg of digest was done on an Ultimate 3000 RSLC Nano system using a 25 cm column (75 μm internal diameter, Reprosil-Pur C18-AQ 1.9 μm phase, PepSep) at a flow rate of 500 nl.min−1 and maintained at 60°C. Solvent A was 0.1% formic acid in LC–MS grade water and solvent B was 0.1% formic acid in 80% ACN. Gradient consisted of a first increase of solvent B from 10 to 30% over 83 min then from 30 to 55% over 17 min. Total run time including column wash and re-equilibration was 120 min.
Tandem mass spectrometry analysis was performed on a Q Exactive mass spectrometer (Thermo Fisher Scientific) equipped with an Easy-Spray nanosource. A full scan was acquired between 350 and 1,400 m/z (mass to charge ratio) at a resolution of 70,000 [automatic gain control (AGC) target of 3.106 ions or 50 ms maximal injection time]. The top 20 precursors were then selected for MS2 analysis at a resolution of 17,500 [AGC target of 1.105 ions, 50 ms maximal injection time, and normalized collision energy (NCE) 27%] with an isolation window of 2 m/z.
Extraction and annotation of MS spectra were performed using Proteowizard (v. 3.0.9992; Chambers et al., 2012), OpenMS (v. 2.4; Röst et al., 2016) software, and BIOTRACS, an in-house solutions developed with MATLAB (R2019). Raw MS spectra were processed using peak picking algorithm to annotate most significant peaks using their m/z ratio and retention time (RT). All peaks below a predetermined background-intensity threshold of 1.104 were removed from data. This threshold was defined according to the mass spectrometer characteristics. The retention time of remaining features were next aligned across all samples to correct potential RT drifts during the chromatography. Peptides were identified by using MASCOT search engine against the Bordetella pertussis Tohama I proteome (proteome ID UP000002676) and regrouped in protein by Fido algorithm (Serang et al., 2010). Trypsin was specified as the enzyme, cleaving after all lysine and arginine residues and allowing up to two missed cleavages. Carbamidomethylation of cysteine (+ 57.021 Da) was specified as fixed modification and oxidation of methionine (+ 15.995 Da) was considered as variable modification. A false discovery rate of less than 5% was applied to both peptide spectral matches and protein based on linear discriminant analysis using a target decoy strategy (Elias and Gygi, 2007).
A filter was applied to keep the features present in more than 80% of samples per group. Subsequently, using BIOTRACS, several QC analyses were performed on processed data mainly to validate homogeneity between samples, considering proteins identified per sample, peptides miss cleavages, samples intensity, and proportion of peptides number used for protein identification. Protein intensities were converted in Log2 scale and normalized by the subtraction of the median intensity of each sample (Dubois et al., 2022).
Lipidomic data generation and analysisLipidomics is defined as the full characterization of lipid molecular species and of their biological roles (Raghunandanan et al., 2019; Bale et al., 2021). Lipids were extracted using Bligh and Dyer liquid extraction method (Bligh and Dyer, 2011; Sündermann et al., 2016). Briefly, 700 μl of the mix chloroform (CHCl3)/methanol (MeOH) at the ratio 1/2 (v/v) were added to pellets. Samples were homogenized by pipetting, transferred in 2 ml vials containing the lysing matrix B (MP Biomedicals) and lysed by bead beating using the Precellys Evolution homogenizer (Bertin Technologies; 3 × 40 s, 6,500 rpm, 4°C, pause of 30 s). Then, samples were incubated in a thermoshaker (Eppendorf) for 10 min at 1,600 rpm and 4°C and centrifuged for 3 min at 10,000 g and 4°C. For each sample, 500 μl of supernatants were isolated. The extraction was repeated two more times with 700 μl of the mix CHCl3/MeOH/H2O at the ratio 1/2/0.8 (v/v), and 2 × 700 μl of supernatants were isolated. Aqueous and organic phases were separated by addition of 500 μl of H2O and CHCl3, mixed by vortexing and then centrifuged for 5 min at 2,500 g at room temperature to accelerate phase separation. Organic phase was isolated (1,000 μl), and 800 μl of CHCl3 was added to repeat organic extraction. Samples were mixed, centrifuged, and organic phase was isolated (800 μl). Then, solvents were evaporated under nitrogen flow at 40°C using the Stuart®evaporator for 1 h. Dried samples were stored at −20°C waiting analysis. As a control, blank samples without biological materials were also prepared using the same preparation protocol.
Lipids were dissolved in 200 μl of isopropanol (IPA) containing an internal standard (IS) and incubated for 10 min at 1,600 rpm and 10°C. This IS is composed of 1 μg.ml−1 of lyso-phosphatidylethanolamine (lyso-PE; 17,0), arachidonic acid (C20:4 D8), PE (C16:0 D31/18:1), phosphatidylglycerol (PG; C16:0 D31/18:1), phosphatidylcholine (PC; C14:0 /14:0 D54), and Ceramide (16,0 D31) in IPA. Then, 25 μl of each sample were pooled to obtain QC samples and the rest of the samples was transferred in HPLC vial inserts for LC–MS analysis. IPA and IPA with IS were used as blanks for LC–MS analysis.
Intracellular lipids were analyzed using an Ultra-High Performance Liquid Chromatography-High-Resolution Mass Spectrometry (UHPLC-HRMS) and a Thermo Scientific Vanquish UHPLC System coupled to a Q Exactive HF mass spectrometer. Three separate injections were performed to acquire spectra in positive and negative ionization modes. Data were acquired in full scan alternating with data dependent acquisition (top 10) to obtain MS/MS spectra.
Lipids were separated on a C18 CSH column (100 mm × 2.1 mm, 1.7 μm, Waters) at 55°C (Isaac et al., 2011). Flow rate was 400 μl.min−1. The mobile phase A consists of ACN/H2O (60/40, v/v) with 10 mM ammonium formate, and 0.1% formic acid and mobile phase B IPA/ACN (90/10, v/v) with 10 mM ammonium formate and 0.1% formic acid. The injection volume was 5 μl. The gradient used for the lipid profiling by UHPLC–MS was performed according to previous study (Isaac et al., 2011).
Q Exactive HF mass spectrometer (Thermo Fisher Scientific) is equipped with HESI-II probe. A full scan was implemented on both positive and negative mode with mass range between 150 and 1,250 m/z (Isaac et al., 2011). The ion source settings were as follows: spray voltage = 3.5 kV for electrospray ionization (ESI) in positive mode and 2.7 kV for negative ESI, capillary temperature = 320°C, sheath gas flow rate = 40, and auxiliary gas = 10 and sweep gas = 0. Data acquisition settings were with AGC (MS1) = 3.106 ions, mass range = 150–2,000 m/z, injection time = 100 ms, and resolution = 120,000. Data acquisition for fragmented sequences was also applied on separated injections, experimental settings were AGC (MS2) = 1.105 ions, mass range = 150–2,000 m/z, injection time = 100 ms, and resolution = 30,000, Top 10, dynamic exclusion = 10 s and NCE = 20–30—40%.
Extraction and annotation of MS spectra were performed using Proteowizard (v. 3.0.9992; Chambers et al., 2012) and OpenMS (v. 2.4; Röst et al., 2016) software, and BIOTRACS. Raw MS spectra were processed using peak picking algorithm to annotate most significant peaks using their m/z ratio and RT. All peaks below a predetermined background-intensity threshold of 1.104 were excluded from data. This threshold was defined according to the mass spectrometer characteristics. The retention times of remaining features were next aligned across all samples to correct potential RT drifts during the chromatography. The resulting feature table was used for QC analysis to correct analytical drifts, in particular intra-batch effects. The coefficient of variation (CV = standard deviation/mean) of features was computed to assess the analytical stability of the features. Features with a CV greater than 30% in QC samples were removed from the data because they were considered as analytically inconsistent. A filter was applied to keep the features present in 80% in the QC samples. Positive and negative mode data were next merged.
Metabolic features were identified using LipidMatch R software (Koelmel et al., 2017). In LipidMatch, a ranking was done on the detected metabolites: Level 1: precursor mass found and confirmed by MS/MS fragments; Level 2: does not apply to our experiment as it refers to all ion fragmentation; Level 3: found class of lipids; Level 4: found precursor mass but unfound MS/MS fragments; and Level 5: unfound precursor mass in the database of LipidMatch. Lipid intensities were converted in Log2 scale and normalized by the subtraction of the median intensity of each sample.
Extracellular metabolomic data generation and analysisSupernatant thawed at room temperature were vortexed and 250 μl of samples were filtered using 0.2 μm centrifugal filter units (VWR; for 5 min at 10,000 g and 4°C). The supernatants were additionally filtered using 10 kDa Molecular weight cut-off (MWCO) centrifugal filter units (VWR; for 15 min at 10,000 g and 4°C). The 45 μl of resulting filtrates were mixed with 90 μl of water and 45 μl of Sodium trimethylsilylpropanesulfonate (DSS) internal standard solution at pH 6, containing phosphate buffer and D2O for the signal lock. The resulting concentration of the DSS was about 0.5 mM. The obtained solutions were vortexed and 155 μl were transferred in 3 mm SampleJet NMR tubes (Bruker®). Finally, tubes were loaded in the 4–6°C pre-cooled SampleJet autosampler before analysis. In addition to samples, blank samples were also prepared using the same preparation protocol, with MilliQ water instead of culture supernatants. The DSS concentration was calibrated with 1.305 mM sodium succinate dibasic hexahydrate solution in triplicates in order to guarantee data accuracy.
The sample analysis was performed using Ascend 600 MHz (Avance III HD) NMR spectrometer (Bruker Biospin) equipped with cryogenically cooled 5 mm QCI (1H/13C/15N/31P) probe head. For each sample a one-dimensional proton acquisition was performed using noesygppr1d pulse sequence, which contain a pre-saturation block for water attenuation during relaxation delay. The spectral width of proton acquisition was 14 ppm using 64 k data points during 3.8 s acquisition time and 4 s relaxation delay. The mixing time delay was 70 ms. Spectra for extracellular metabolites contained 512 scans for about 70 min of total acquisition time. The free induction decay (FID) raw data were processed by Fourier transformation using 0.3 Hz exponential apodization function within 64 k data points. The resulting spectra were phase and baseline corrected. The spectra alignment was done according to internal standard DSS chemical shift. Finally, the metabolites quantification was performed using Chenomx NMR suite 8.3. The metabolites concentrations were reported in mM and exported in Excel format file for statistical analysis.
Biostatistics analysesEach omics dataset was processed independently through a unique statistical pipeline consisting of three steps: QCs, supervised univariate analysis, and longitudinal clustering. An additional pathway enrichment step was also performed on transcriptomic and proteomic data.
For QCs, principal component analyses (PCA) and hierarchical clustering (based on Ward distance) were used to measure biological variability and ensure samples were more similar within (technical and biological replicates) than across time points.
A variance partition step was also conducted to investigate the relative contribution of time, technical variables (sequencing batch, bioreactor), and noise on omics measurements. This step was carried out using the Gaussian Processes R package lgpr (Timonen et al., 2021).
Univariate analysis was done using both (moderated) t-test and Gaussian Processes to quantify the effect of time on expression data and measure differences between time points. A linear mixed model was fitted on transcriptomic and proteomic data using Limma (combined with Voom in the former case; Law et al., 2014) to account for the correlation between samples from the same bioreactor. The time and batch effects were, on the other hand, included in the model as categorical fixed effects. For each gene/protein, a moderated t- and F-statistics were computed to test whether pairwise differences among time points (contrasts) were equal to zero separately and altogether. F-statistics were used as a measure of significance of the time effect along the whole trajectory. To identify major variations, the significativity threshold for gene/protein variations was fixed to |log2FC| > 1 with an adjusted p-value < 0.05. To identify fine changes of specific genes transcription or proteins syntheses, variations with a |log2FC| > 0.5 and an adjusted p-value < 0.05 were scrutinized. For other omics than transcriptomic and proteomic, a paired t-test was used in place of Limma.
Unlike linear mixed model, Gaussian processes are non-parametric approaches that model more complex covariance structures and detect non-linear effects of both categorical and continuous covariates (as well as their interactions). The lgpr package provides a measure of covariate effects (proportion of explained variance) for each omics feature as well as a measure of covariate relevance. Using the same three effects (time, sequencing batch and bioreactor), we found that the percentage of explained variance returned by lgpr led to an almost identical gene ranking as linear mixed model F-statistics.
A longitudinal clustering was then carried out using Dirichlet process Gaussian process mixture model (DPGP; McDowell et al., 2018) that combines a Dirichlet process to determine the number of clusters and a Gaussian process to model the trajectory along the time. Both approaches are based on nonparametric methods. For each omic, only the features with an explained variance larger than 25% were fed into the model, threshold set to filter out non-informative features. The 25% value was found to correspond to the 5% adjusted p-value in the Limma-lgpr comparison on transcriptomic data. DPGP returned clusters of omics features presenting shared trajectories along the time.
A last step of pathway enrichment was performed on transcript and protein clusters, assuming that features with similar trajectories shared biological functions. A standard over-representation analysis based on the Fisher’s exact test (Huang et al., 2009) was carried out on Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) databases using clusterProfiler R package (Yu et al., 2012).
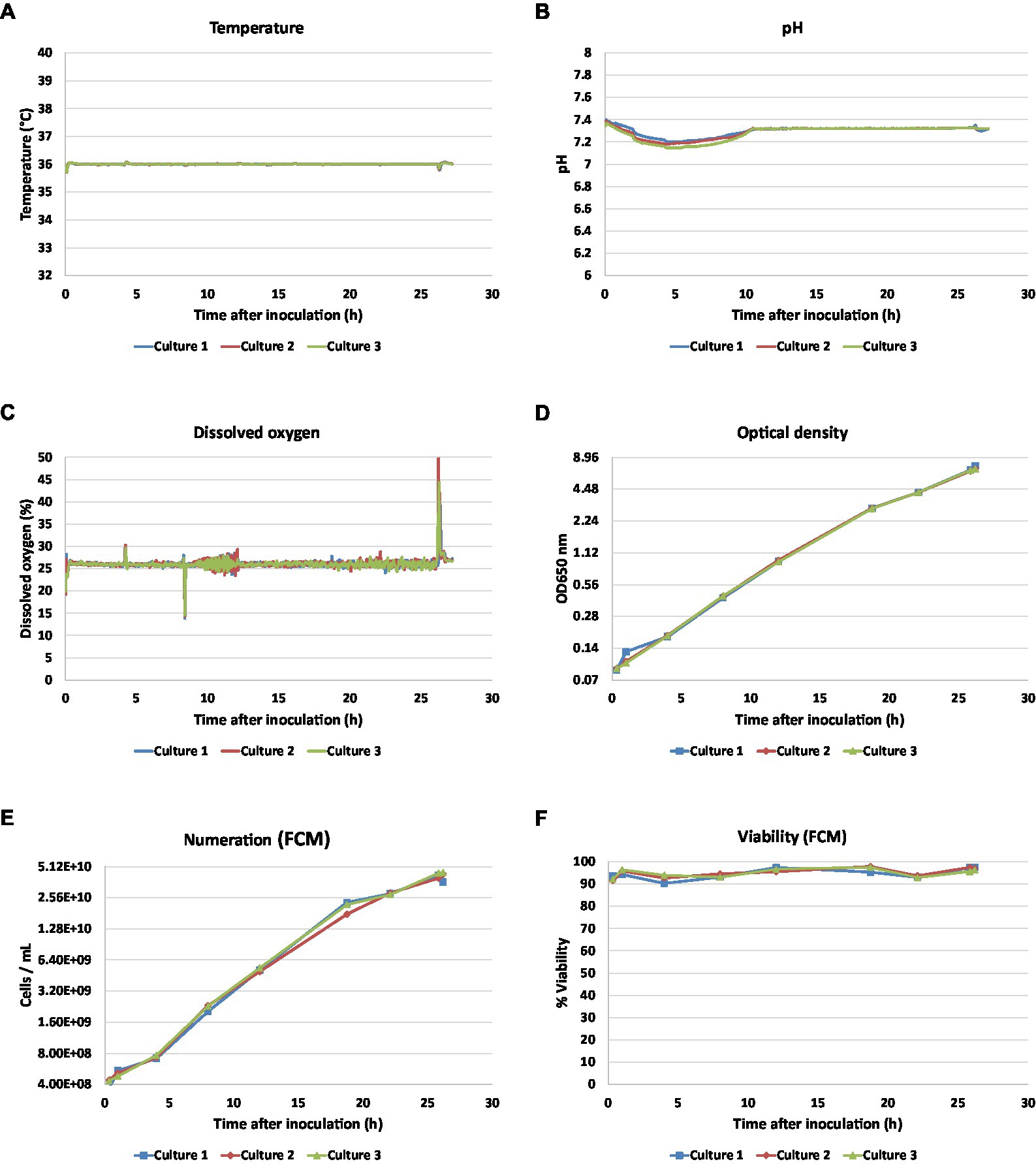
Results Standardized growth and biosynthesis of vaccine antigens through experimental culturesTo mimic industrial culture conditions, three independent culture batches were performed in hydrolysate casein-like (HC-like) chemically defined medium (Izac et al., 2015). As expected, no significance difference in temperature, pH, and dissolved oxygen (DO) were observed throughout cultures (Figures 2A–C). Similar and minor perturbations were observed regarding pH regulation at the beginning of the culture and dissolved oxygen fluctuation linked to sampling in the three fermenters. Bacterial growth and viability were monitored by OD650 measurements and flow cytometry analyses (Figures 2D–F). Independent culture replicates showed similar growth and a viability higher than 90% was observed throughout cultures. The end of culture was identified by a DO peak and a CER drop at 26 h. Two main growth phases could be distinguished during culture: the first, between 0 h and 18 h 45 min (early to middle logarithmic phase), was characterized by an OD and a numeration growth rate of 0.19 and 0.21 h−1 respectively, the second, between 18 h 45 min and 26 h (mid to late log phase), differed by lower growth rate (0.12 and 0.09 h−1, for OD and numeration respectively).
Figure 2. Monitoring profiles of three independent bioreactor cultures of Bordetella pertussis Tohama I. Temperature (A), pH (B), DO (C) were monitored inline by Ambr system, OD650(D), bacteria numeration by flow cytometry (E), and viability measured by flow cytometry (F) were offline determined on 8 time points.
Final amounts of PT, PRN, Fim2, and FHA were measured from whole bacterial suspensions using ELISA. The mean antigen concentrations were 10.5 μg.ml−1 (CV = 3.0%) for PT, 24.2 μg.ml- 1 (CV = 4.6%) for PRN, 211.6 ng.ml−1 (CV = 2.7%) for Fim2, and 150.4 μg.ml−1 (CV = 1.8%) for FHA. Based on the small coefficient of variations observed across batches, we thus extrapolated that our experimental biological triplicates were similar.
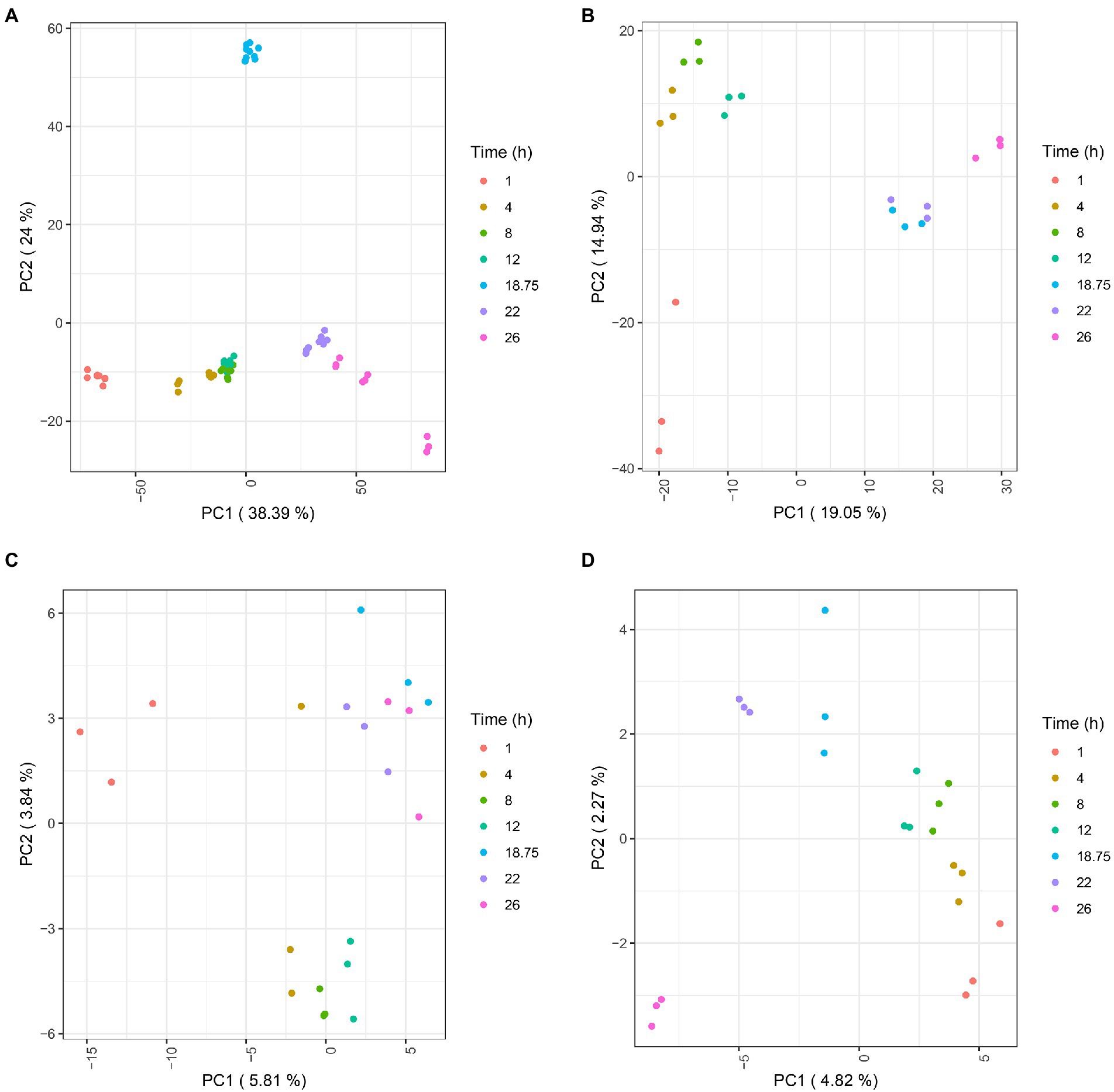
Longitudinal multi-omics reveals a global metabolic switch during Bordetella pertussis cultureTranscripts, proteins, lipids, and metabolites were analyzed from three independent batch cultures at different times points (T = 1 h, 4 h, 8 h, 12 h, 18 h 45 min, 22 h and 26 h; Figure 1). Following raw data processing, a total of 3,581 transcripts, 1,340 proteins, 99 lipids, and 37 extracellular metabolites were identified and quantified (Supplementary Tables 1–4). PCA was conducted on each omics dataset separately to visually check the consistency between the biological replicates and to evaluate the relative contributions of experimental factors on data variation (Figure 3). PCA within each omics revealed that samples were clustered according to time points, reflecting a high homogeneity across biological replicates and therefore, high reproducibility in all omics. As expected from the literature (Metz et al., 2017; Dienstbier et al., 2019), intra time point variation was higher in proteomic and lipidomic data due to the larger technical variability and weaker variation of lipid and protein amounts during culture compared to transcripts. Due to technical sampling issues that were later confirmed by the PCA analysis, proteomic and lipidomic samples collected at T = 1 h were excluded from downstream analyses. After data quality assessment, statistical longitudinal analyses were performed.
Figure 3. PCA plots of transcriptomic (A), proteomic (B), lipidomic (C), and extracellular metabolomic (D) data. For transcriptomic, each single dot represents a library replicate. For other omics, each single dot represents a biological replicate.
We first aimed at identifying the feature variations that were the most associated with time using Longitudinal Gaussian Processes (LGPR; Timonen et al., 2021; Supplementary Tables 1–4). Gaussian processes are flexible non-parametric approaches, able to model complex non-linear and non-stationary signals. In agreement with initial PCA, proteomic and lipidomic data showed higher technical variations than transcriptomic and extracellular metabolomic data. Features with more than 25% of their variance explained by the time were considered of interest in downstream analyses. This threshold was found to correspond approximately to a 5% q-value in a linear mixed model (see Material and Methods).
Overall, 3,080 out of 3,581 transcripts, 723 out of 1,340 proteins, 42 out of 99 lipids, and 33 out of 37 extracellular metabolites passed this cut-off.
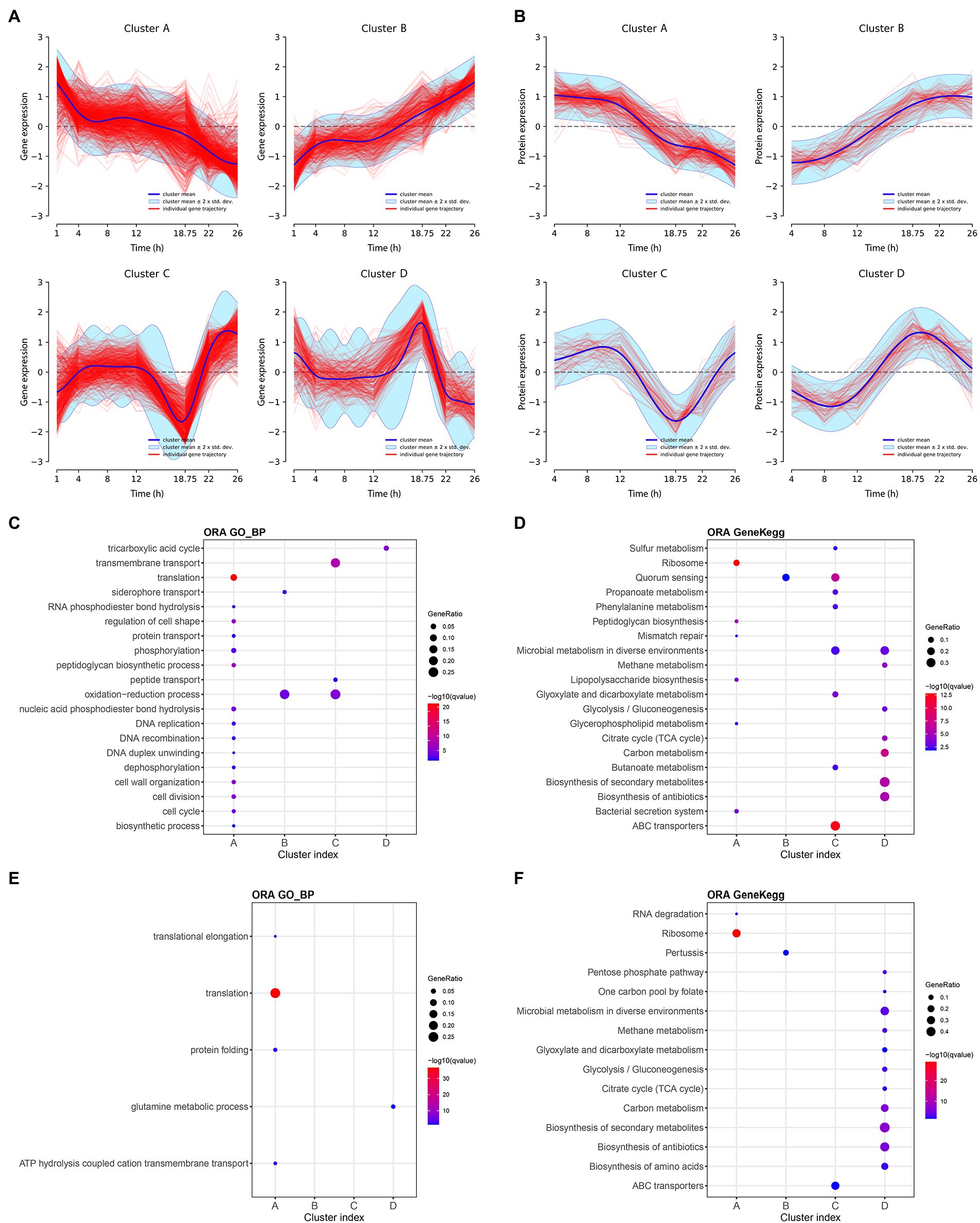
Using the features selected above, we subsequently investigated similarities among feature trajectories using longitudinal clustering in each omics dataset (McDowell et al., 2018). Cluster analyses identified 13 and 11 clusters in transcriptomic and proteomic data, respectively (Supplementary Figures 1, 2). Next, clusters with similar profiles were merged and regrouped in larger clusters named A–D for both transcriptomic and proteomic data (Supplementary Tables 1, 2). For the ease of graphic representation, representative profiles of these large clusters are depicted in Figures 4A,B.
Figure 4. Main longitudinal clusters trajectories and pathway enrichment results of transcriptomic and proteomic data of Bordetella pertussis culture. Main longitudinal clusters trajectories of transcriptomic (A) and proteomic (B) data. X-axis correspond to the time and Y-axis correspond to the gene expression or the protein intensity. Blue line corresponds to the cluster mean, light blue interval corresponds to the cluster mean ± 2 × SD and red line corresponds to the individual transcript or protein trajectories. Over representation analysis (ORA) pathways enrichment on Gene Ontology and KEGG databases for transcriptomic (C,D, respectively) and proteomic (E,F, respectively). Enriched pathways are shown in Y-axis and associated cluster in X-axis. Colors of spots (blue to red) are according to the significance of pathways enrichment in each cluster. Size of spots are according to the Gene ratio (= number of transcripts or proteins in a cluster which mapped on a pathway/Number of transcripts or protein in this cluster which mapped in the database). Maximum number of pathways = 20. Maximum q-value = 0.05.
All clusters trajectories are globally monotonous up to 12 h of culture. After 12 h, the profiles of clusters A (1,132 transcripts and 260 proteins) decrease while the profiles of clusters B (657 transcripts and 81 proteins) increase until the end of culture. Clusters C are composed of 471 transcripts and 88 proteins and decreases over 12 h, before going up after 18 h 45 min until the end of culture. Conversely, the profile of clusters D (391 transcripts and 130 proteins) increases after 12 h, up to 18 h 45 min before decreasing until the end of culture. The consistency between proteins and transcripts clusters was evaluated by measuring the percentage of proteins from a cluster profile found within their matching transcripts in the same cluster profile. The consistency between proteins and transcripts was 70, 51, 65, and 52%; for the clusters A–D, respectively. Moreover, between 12 h and 18 h 45 min, 1,790 genes (177 up- and 309 down-regulated, |log2FC| > 1) and 399 proteins (17 up-, including five ON proteins, and 17 down-regulated, including eight OFF proteins, |log2FC | > 1) are differentially expressed (adj p-value < 0.05; Supplementary Tables 5, 6). Between 18 h 45 min and 22 h, 1,671 genes (310 up and 91 down regulated) and 90 proteins (five up including three ON proteins and zero down regulated) were differentially expressed (adj value of p < 0.05). Finally, clustering reveals a specific event at 18 h 45 min of culture with irreversible impact highlighted by both clusters A and B. Both clusters C and D indicate a steep change of omics profiles before returning to their respective general paths of expression suggesting a transient physiological state.
To functionally characterize clusters, pathway enrichment analyses (Huang et al., 2009) were next performed with the groups of transcripts/proteins that constitute each cluster using the KEGG and GO databases (Supplementary Tables 7–10). For each database, the top 20 pathways (ranked by q value, q values <0.05) identified with the transcriptomic and proteomic data of the clusters are, respectively, reported in the Figures 4C,D. The cluster A, both in transcriptomic and proteomic, is mainly associated with several biosynthesis pathways, such as ribosome, translation, peptidoglycan biosynthesis, and lipopolysaccharide biosynthesis pathways. Over time, after 12 h of culture, pathways of the cluster A were less used by B. pertussis. This finding is consistent with the observation of a growth rate reduction. The decrease of such biosynthesis pathways was previously highlighted during the transition from exponential to stationary phase as well as under glutamate limitation (Nakamura et al., 2006). Cluster C enrichment revealed that the expression of 55 ABC transporter genes and the synthesis of 15 ABC transporter proteins decrease specifically at 18 h 45 min of culture. Cluster D was enriched with transcripts and proteins involved in TCA cycle and glycolysis/gluconeogenesis pathways.
The decrease of multiple biosynthesis pathways and the increase of ABC transporter genes after 18 h 45 min point toward one or several putative culture medium components starvation.
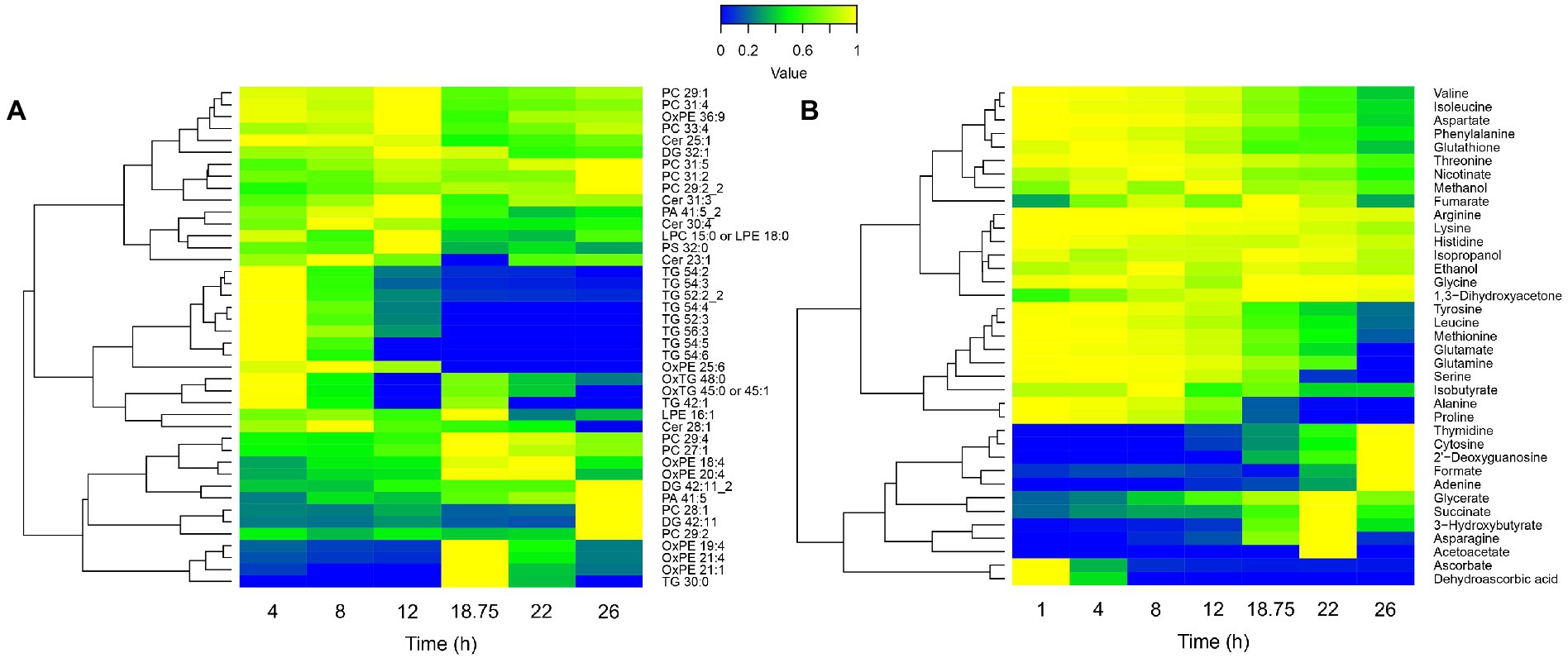
To confirm this hypothesis and further decipher the physiology behind the breakpoint occurring at 18 h 45 min, the longitudinal lipidome and extracellular metabolome of Tohama I were investigated (Figures 5A,B). The lipidomic analyses showed different trends of lipid variation across the lipidome (Figure 5A). Phosphatidylcholine (PC) mostly accumulated at the end of the culture. Oxydated phosphatidylethanolamine (OxPE) increased at 18 h 45 min, which suggests an occurring oxidative stress. Conversely, triglycerides (TG) were totally degraded at 18 h 45 min. Extracellular metabolomics analyses showed large variations of metabolite concentrations in the culture medium along the culture process (Figure 5B). In agreement with previous studies performed in modified SS-medium, purine (adenine), pyrimidine (cytosine) and nucleosides (deoxyguanosine and thymidine) accumulated at the end of the bioreactor culture (Branco dos Santos et al., 2017). In addition, several other metabolites, such as acetoacetate, hydroxybutyrate, and succinate also accumulated from 18 h 45 min to 22 h.
Figure 5. Lipidomic (A) and extracellular metabolomic (B) longitudinal data heatmaps of B. pertussis culture. Lipid intensities and metabolite concentrations are represented from 0 (blue) to 1 (yellow), 1 representing the maximum intensity/concentration of every feature. Features are clustered according to their trajectory similarities throughout the culture. Cer, ceramide; DG, diglycerides; LPC, lysophosphatidylcholine; LPE, lysophosphatidylethanolamine; Ox, oxydated; PA, phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PS, phosphatidylserine; and TG, triglycerides.
Diverse carbon sources consumption could be noticed. Among those carbon sources, alanine and proline were completely consumed between 18 h 45 min and 22 h (Figure 5B). Proline is known to be one of the main carbon sources in modified SS-medium (Stainer and Scholte, 1970; Izac et al., 2015) with glutamate. Thereby, we hypothesized that the observed growth rate decrease, the biosynthesis pathways slowdown, the energy storage consumption, and the marked modulation at 18 h 45 min revealed by longitudinal clustering analysis may be the consequences of a proline starvation. To validate this hypothesis, proline and glutamate metabolisms were next investigated.
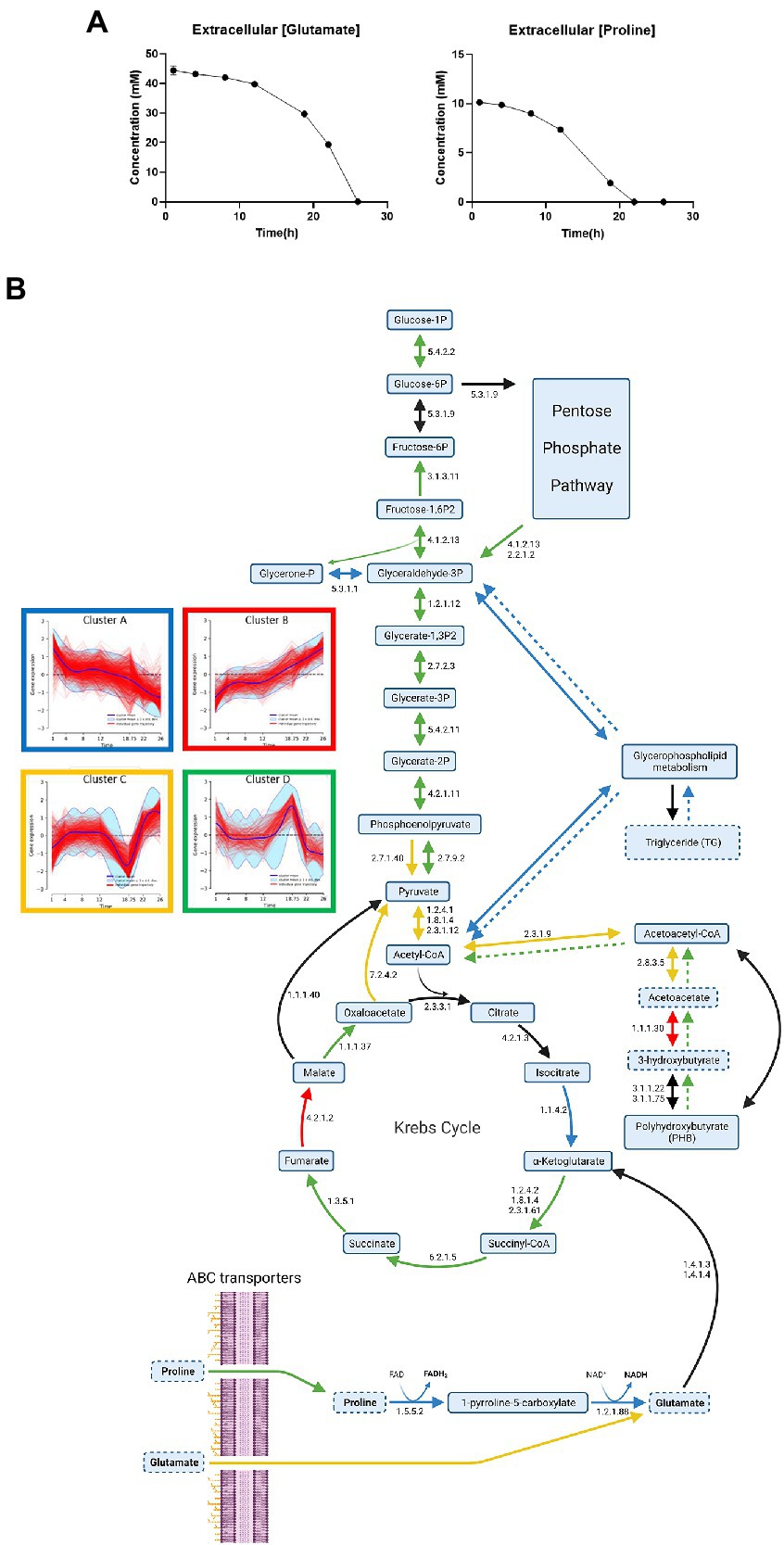
Proline starvation induces transient metabolism with energy stock consumption and impacts central metabolismThe concentrations of glutamate and proline were quantified in the culture medium along the culture (Figure 6A). Extracellular sources of proline and glutamate were fully consumed after 22 and 26 h of culture, respectively. Thus, we can assume that a proline starvation occurs between 18 h 45 min and 22 h, while 44% of the initial extracellular glutamate is still available after 22 h of culture. Therefore, we hypothesized that a decrease of proline catabolism followed by a compensatory increase of glutamate uptake should simultaneously occur between 18 h 45 min and 22 h of culture. To verify this hypothesis, proline and glutamate metabolisms were investigated through an analysis of transcriptomic (Figure 6B) and proteomic data.
Figure 6. Impact of proline starvation on central metabolism during Bordetella pertussis Tohama I culture. (A) Longitudinal evolution of glutamate and proline extracellular concentrations monitored by NMR analyses. (B) Central metabolism gene expression profiles during B. pertussis Tohama I culture. Blue, red, yellow, and green arrows show the enzymes encoded by genes of the clusters A–D (Figure 4A), respectively. Black arrows indicate a discrepancy of cluster between the different enzyme genes. Numbers next to arrow reactions are the EC numbers of the reaction. Dashed arrows are reactions deducted from lipidomic or metabolomic results using the same color code than transcriptomics data. Lipids/Metabolites with dashed outlines were quantified in lipidomic or extracellular metabolomic analyses. This figure was created with BioRender.com.
On one hand, proline ABC transporter genes (bp2055, bp2056, and bp2057) expression decreased (log2FC = −1.5) between 1 and 4 h. The expression of these genes remained stable between 4 and 12 h, increased from 12 h to 18 h 45 min, and then decreased from 18 h 45 min to 26 h of culture (Kyoto Encyclopedia of Genes and Genomes, 2022; Supplementary Table 1). Following a 22 h preculture, B. pertussis bacteria have consumed the stock of proline available from the preculture medium and are on starvation. The high expression of proline transporter genes at 1 h could thus be explained by the exposure to the high proline concentration of the fresh culture medium which triggers the expression of proline uptake genes.
Following its uptake, proline is known to be metabolized in glutamate using two enzymatic reactions (EC 1.5.5.2 and 1.2.1.88), producing 1 FADH2 and 1 NADH, mainly catalyzed by PutA (proline dehydrogenase, BP2749; Surber and Maloy, 1998). Then glutamate may be converted into 2-oxoglutarate and joins TCA cycle and central metabolism (Figure 6B). This is consistent with the sudden decrease of the putA gene (cluster A) expression (Log2FC = −4.7) and of the PutA protein (cluster A) synthesis (Log2FC = −1) between 12 and 22 h (Supplementary Tables 5, 6). Taken together, these results indicate that, proline uptake decreased after 18 h 45 min and the activity of the main proline catabolism pathway decreased after 12 h, in agreement with the proline starvation observed between 18 h 45 min and 22 h.
On the other hand, glutamate ABC transporter genes (gltJ = bp0057, gltK = bp0055/BP0767, gktJ = bp0056, and gltL = bp0768) expression levels were globally stable until 18 h 45 min, and then increased until the end of the culture (cluster C; Kyoto Encyclopedia of Genes and Genomes, 2022). The expression levels of other glutamate transporter genes such as gltS, encoding a sodium/glutamate symporter, increased between 12 and 22 h (log2FC = +0.8). Similarly, while the expression of bp3831, encoding a putative glutamate transporter (Keidel et al., 2018) decreased between 12 h and 18 h 45 min (Log2FC = −1.8), a marked expression increase of this gene was also observed between 18 h 45 min and 22 h (Log2FC = +2.0), (Supplementary Tables 5, 6). Those results show that transcriptomic and proteomic data are consistent with extracellular metabolomic data and confirm that glutamate uptake increases after proline starvation.
Proline metabolization is a major source of energy. Thus, a starvation of this amino acid conducts to a loss of energy which may trigger energy internal stock consumption. Proline is metabolized by TCA cycle via glutamate and poly-β-hydroxybutyrate (PHB) is connected to central metabolism via acetyl-CoenzymeA (acetyl-CoA). Thereby, the expression of genes and the synthesis of proteins involved in the central metabolism were investigated (Figure 6B). In B. pertussis, the TCA cycle was recently demonstrated to be entirely functional in the Tohama I strain (Izac et al., 2015; Branco dos Santos et al., 2017; Gonyar et al., 2019). In the present study, the expression of most TCA cycle genes and the synthesis of corresponding proteins increased at 18 h 45 min of culture (cluster D; Figure 6B; Supplementary Tables 1, 2). Transcripts and proteins involved in the conversion of citrate to isocitrate and 2-oxoglutarate (EC 4.2.1.3 and 1.1.1.42) decreased after 12 h of culture. Similarly to a large part of the TCA cycle, determinants of gluconeogenesis activity increased at 18 h 45 min of culture (cluster D). In B. pertussis, gluconeogenesis is necessary to supply carbon to several pathways including the pentose phosphate pathway (PPP) and the glycerophospholipid metabolism (Figure 6B). PPP is used to synthetized DNA precursors (Kyoto Encyclopedia of Genes and Genomes, 2022). Thus, the reduced growth and the decrease of several biosynthetic activities are consistent with the slowdown of central metabolism after 18 h 45 min of culture. The expression of most of glycerophospholipid metabolism genes decreased at 18 h 45 min (cluster A). As glycerophospholipid metabolism allows to synthetize triglycerides, this observation is in agreement with the absence of triglyceride at 18 h 45 min (Kyoto Encyclopedia of Genes and Genomes, 2022).
Taken together, these findings strongly suggest a proline starvation at 18 h 45 min of culture in modified SS-medium leading to a major modulation on B. pertussis central metabolism. As modified SS-medium is a commonly used medium for B. pertussis cultivation (Fyson et al., 2017), in laboratory and industrial processes, the impact of this metabolic switch was further assessed on antigen production and virulence.
Bordetella pertussis antigen production and global virulence regulation during cultureAntigens used in aP vaccines are B. pertussis virulence factors. To document major antigens production and evaluate the impact of proline starvation on their production, a longitudinal antigen quantification was carried out (Figure 7).
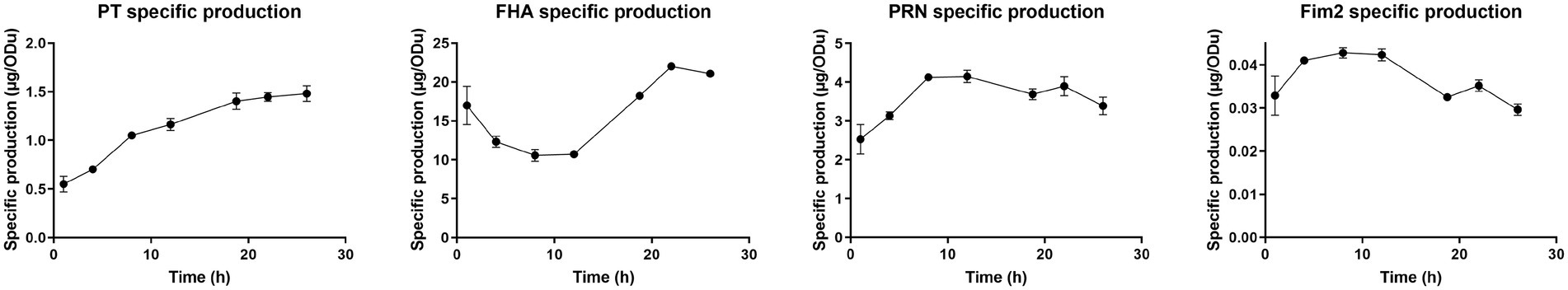
Figure 7. Specific antigen production throughout Bordetella pertussis culture. ELISA antigen quantification was performed for each antigen on lysed crude harvest and normalized using biomass measured with OD650.
The specific total production of PT increased by 3-fold (0.55–1.48 μg.ODu−1) from the beginning to the end of culture, with a marked slowdown of after 18 h 45 min. Indeed, specific production speed exhibits non-significant 3-fold decrease after 18 h 45 min compared to the 12–18 h 45 min interval. The specific total FHA production was stable between 4 and 12 h and then sharply increased until 22 h by 2-fold (10.73–22.00 μg.ODu−1). The specific total PRN and Fim2 production increased from 1 to 8 h after inoculation, remained stable from 8 to 12 h and then slightly decreased until the end of culture (between 12 and 26 h).
Transcriptomic and proteomic analyses were used to monitor transcripts and proteins involved in aP vaccine antigen production throughout culture (Supplementary Tables 1, 2, 5, 6). Data showed that ptx operon transcription increased throughout culture (log2FC ≈ + 1 between the beginning and the end of culture) and Ptx subunits amounts slightly increased until 18 h 45 min of culture (log2FC ≈ +0.5 between 4 h and 18 h 45 min) and then remained stable (Supplementary Figure 3). PT is folded by DsbAC in the periplasm and externalized by Ptl type IV secretion system (Stenson and Weiss, 2002; Stenson et al., 2003). The transcription of dsbA increased from 4 h to 18 h 45 min (log2FC = +1.2), then decreased until 22 h (log2FC = −0.6 between 18 h 45 min and 22 h) and increased until the end of the culture (log2FC = +0.7 between 22 and 26 h). The amount of dsbC transcripts and DsbA/DsbC concentrations were stable throughout the culture. Ptl operon expression and the corresponding protein concentrations remained stable (Supplementary Figure 4). The concentration of FhaB, the precursor of FHA, slightly varied during culture. Conversely, fhaB expression increased by almost two-fold (log2FC = +0.9) between 4 h and 18 h 45 min of culture. FhaB is transported through the outer membrane by FhaC, processed by CtpA and released by SphB1 (Nash and Cotter, 2019). Transcripts of fhaC increased from 8 h to 18 h 45 min (log2FC = +0.5) and then decreased between 18 h 45 min and 22 h (log2FC = +0.9). The transcriptions of sphB1 and ctpA were stable throughout the culture. SphB1 quantity slightly increased between 4 and 22 h (log2FC = +0.6) and FhaC and CtpA concentrations remained stable. Prn gene expression increased until 12 h and then decreased until the end of the culture (log2FC = −0.8 between 12 and 26 h). Similar observation was observed for PRN in proteomic but with lower variations. Fim2 antigen is composed of Fim2 and FimD subunits (Scheller and Cotter, 2015). Fim2 gene expression was stable during the culture and fimD expression increased from 8 h to 18 h 45 min (log2FC = + 0.6) and decreased between 18 h 45 min and 22 h (log2FC = −0.8). Fim2 concentration was stable while FimD decreased from 18 h 45 min to 26 h (log2FC = −0.5). Fim2 antigens is folded by the FimB chaperone and externalized by FimC usher proteins (Scheller and Cotter, 2015). Gene expression of fimBC was stable from 4 h to 18 h 45 min and decreased between 18 h 45 min and 22 h (log2FC = −0.5-0.7). The amount of FimB was stable throughout culture while FimC concentration increased from 8 to 22 h (log2FC = +0.6). The transcription of fim3 was stable throughout culture. Fim3 protein was not detected by our proteomic analysis. In agreement with ELISA data, those results suggest that dsbA, fhaC, prn, and fimD gene expressions and Ptx, Prn, and FimD protein productions might be impacted by proline starvation.
To determine whether the proline starvation is concomitant to a global virulence modulation of B. pertussis, the master virulence regulon BvgASR profile was analyzed (Figure 8). BvgAS is a two-component system of B. pertussis that regulates the expression of virulence activated genes (vags). Among those genes, bvgR encodes for a phosphodiesterase, which turns off the expression of virulence repressed genes (vrgs; Chen and Stibitz, 2019). Here, bvgA increased from the beginning of the culture until 12 h, remained stable until 18 h 45 min, and then decreased until the end of the culture (log2FC = −0.8 between 18 h 45 min and 26 h). The bvgS gene was stable up to 18 h 45 min and then decreased until the end of the culture (log2FC = −0.7 between 18 h 45 min and 26 h) while bvgR, was stable up to 12 h and then decreased until the end of the culture (log2FC = −0.7 between 12 and 26 h). BvgAS protein amounts increased slightly (log2FC ≈ +0.5) between 4 and 22 h while BvgR remained stable. Those results suggest a moderate down regulation of bvgASR genes after 18 h 45 min. RisAK is a two-component system of B. pertussis involved in the regulation of virulence repressed genes (vrgs). In contrast to bvgASR, risAK transcripts and RisA proteins were stable throughout the culture. As RisK was not detected in two out of three biological replicates at 18 h 45 min, it was not possible to conclude on its variation. Finally, those data indicate tha
留言 (0)