
記住我


Biofilm-based microbial infections have become a severe threat to public health. Based on the increasing cases of polymicrobial biofilm infections, this review article explores the recent updates on polymicrobial biofilm pathogens, interactions between and within microorganisms that exacerbate biofilm formation, and model systems to study polymicrobial biofilms. We have also explored the metagenomic approaches towards the surveillance of polymicrobial biofilms and the recent trends to mitigate polymicrobial biofilms, such as quorum sensing inhibitors, nanoparticle- and nanoconjugate-mediated therapy, antimicrobial photodynamic therapy (aPDT), phage therapy, combinatorial probiotics, etc.
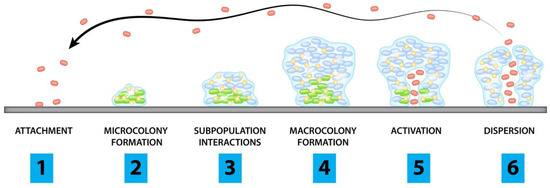
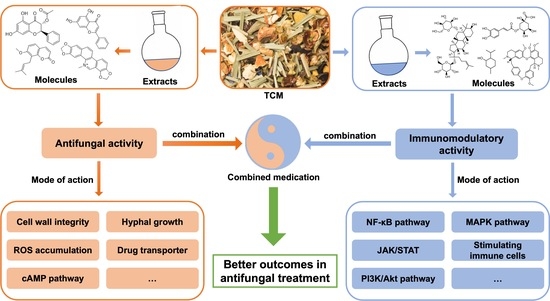
3. Polymicrobial InfectionsThe colonisation of one microorganism can influence the colonisation of the other microorganisms in the host. In respiratory tract infections, viruses promote bacterial infection by suppressing the immune system, destroying epithelial cells, and upregulating the expression of molecules essential for the adhesion of bacteria. These viruses are also involved in middle-ear infections caused by bacteria, resulting in otitis media [18]. Polymicrobial biofilm infections affecting different parts of the body are depicted in Figure 1. Dental caries is the most common dental infection, affecting almost 60 to 80% of children and adults. The tooth surface serves as a site for the attachment of conditioning film (made up of salivary proteins and carbohydrates), to which the microorganisms form the biofilm [7]. Studies reported that approximately 48 h are required to transform individual cells into biofilms. S. mutans, the most common etiological agent causing dental caries, primarily attaches via glucosyltransferases and initiates the coaggregation of other bacteria, resulting in a polymicrobial biofilm. Such a type of biofilm prevents the host immune response by ceasing the host-signaling pathways [19]. Otitis Media or middle-ear infection affects the Eustachian tube located between the tympanic membrane and the inner ear. It is a childhood disease and rarely incident with the death of the person. When the infection is severe, it results in the loss of hearing ability of the patients. The bacteria which cause middle-ear infections are the commensals such as S. pneumoniae, H. influenza, Moraxella catarrhalis, which, in association with viruses such as influenza A virus, Adenovirus, Rhinovirus and Respiratory Syncytial Virus (RSV), form the infection. Monomicrobial infections caused by these microbial species have significant effects, but they also predispose the host to polymicrobial infections [7]. Lung infection, especially Cystic Fibrosis (CF), has been extensively studied for polymicrobial infection. It is an autosomal recessive disorder associated with microbial infection and leads to respiratory failure. Due to the lack of mucociliary clearance in CF patients, the inability to clear the pathogens trapped in the airways causes chronic polymicrobial infections. Pathogens such as P. aeruginosa, S. aureus, Streptococcus milleri group, Burkholderia cepacia, Stenotrophomonas maltophilia, H. influenzae and C. albicans cause infection in the airways. It was reported that the initial colonizers of lung infections in CF are S. aureus or H. influenzae, and the later colonizers are P. aeruginosa and finally B. cepacia [7,20]. Wound infections are formed when a patient’s epithelial barrier is compromised due to underlying conditions such as obesity and diabetic mellitus. Biofilm formed by the microbes delays the healing of the wound and renders it resistant to antibiotic treatment. Bacterial species involved in chronic wound infection are P. aeruginosa, Enterococcus sp., S. aureus, Streptococcus sp., and E. coli. Studies demonstrated that co-infection with Bacteriodes fragilis and E. coli showed polymicrobial interaction among both species, resulting in inflammation and pus-containing wounds [20]. Understanding the microbial species involved, predisposing factors of the disease progression, and the polymicrobial interaction between microorganisms are essential for diagnosing and developing treatment strategies. 4. Polymicrobial InteractionsMicrobial interactions play a vital part in maintaining microbiome structures. Mutualistic or commensal relationships among microbes develop into positive interactions, whereas parasitic or pathogenic interactions are negative. Novel techniques have helped to explore and characterize the microbial interactions occurring in the microbiome and enable the manipulation of the microbiome for better and improved medical treatments, environmental, and agricultural applications [21]. Most of the biofilms are observed as polymicrobial biofilm communities attached to inert or living surfaces. Polymicrobial biofilms are a group of various microorganisms, such as bacteria, fungi, and viruses, that live in a coordinated manner. In this, bacterial populations are embedded in the exopolysaccharide (EPS) matrix produced during their developmental stage [18]. The exopolysaccharide matrix consists of exopolysaccharides, extracellular DNA, lipids, and proteins. The EPS matrix is responsible for significant structural and functional properties of biofilms, including their survival and virulence [15].Interestingly, bacterial biofilms formed during infections develop into polymicrobial interactions. The biofilm bacteria recruit other bacterial species to pursue polymicrobial interactions and regulate the gene pool provided by each of them. Thus, biofilm as a whole works synchronously to control and regulate the survival mechanisms of individual members. The requirements of biofilms, such as attachment to the target site, stimulation of host cellular senescence mechanisms to prevent the shedding of bacteria, and the production of plasma exudate for nutrition through local inflammation, are carried out by the gene pool of biofilm. The advantage of having polymicrobial groups in biofilm is that the genes contributed by each of the individual colonies perform their basic requirements [22,23,24]. The advantage of genomic plurality in polymicrobial biofilms is that it allows the development of novel strains and fosters persistent infections. The supragenome of some species in biofilms is shared among the other community members, developing its total gene pool. The genomic plurality is regulated by horizontal gene transfer occurring in the biofilm communities. The mechanisms other than the gene pool that enable their survival are passive resistance, quorum sensing pathways, metabolic co-operation, and by-product influence [12]. Polymicrobial interactions are generally observed on the host and environment surfaces. Though they are present in different body sites, the most studied and diverse polymicrobial interactions are from the oral cavity. The interactions among polymicrobial communities involved in pathogenesis are complex. The interactions may cause tight competition for nutrients and space or develop cooperative relationships to enable the growth of each partner in the polymicrobial colony [10]. The polymicrobial interactions can be synergistic, additive, or microbial interference mechanisms. In synergistic associations, establishing one microbe in a particular niche favors the host for the entry and colonization by one or more pathogens causing infections or disease [25]. More severe disease symptoms are often observed during synergy than the occurrence of conditions by the individual pathogens alone. For instance, studies showed wide microbial diversity during human periodontitis infections. The microbial diversity observed at the subgingival crevice indicated synergistic interactions. Likewise, highly dense microbial species were found in the lungs of CF patients associated with polymicrobial infections. The severity of lung infection is affected by the pathogens, S. aureus, H. influenzae, P. aeruginosa, and B. cepacia [26].Additive polymicrobial interactions are observed within the biofilms attached to natural or artificial surfaces in the human system. Additive interactions lead to infections such as bacteremia, liver and soft tissue infections, otitis media, brain, lung, and abdominal abscesses by the combined effect of two or more non-pathogenic microorganisms [18]. The oral plaque biofilms are formed by several aerobic and anaerobic Gram-negative and positive bacteria and some Candida sp. They cause additive periodontal diseases. Actinomyces and Streptococci sp. initially colonize the tooth surface, and their interactions lead to the co-aggregation and co-adhesion of other pathogens [27]. In microbial interference, polymicrobial interactions between potent pathogens or probiotic organisms and pathogens may develop a niche unfavorable for colonization by other organisms [28]. For example, a flavivirus, GBV-C, resembles the hepatitis C virus but is not pathogenic in humans. GB virus C replication takes place in lymphocytes. The in vitro viral infections are associated with a decreased mortality rate in HIV-infected persons. The reduced mortality is due to the inability of HIV to replicate in the lymphocytes infected with GBV-C [28]. Antimicrobial synergism is observed with polymicrobial biofilms where the infectious agent shows higher antibiotic susceptibility than the individual species. For instance, in antimicrobial susceptibility testing, 60% of planktonic cells were inhibited with antibiotic combinations. In contrast, susceptibility towards antibiotics was observed in only 22% of bacteria in their biofilm state [29]. 4.1. Bacterial-Bacterial BiofilmsBacteria exhibit co-aggregation and co-localization mechanisms to interact with their partner species within a biofilm. Beneficial partner species with different genes can be selected within the biofilm by reversible co-aggregation methods. In contrast, during the co-localization process, favorable growth conditions are provided by beneficial bacteria to enable biofilm development [30]. The initial colonization of one species helps other partner bacteria. The initial colonizer prepares the substrate surface on which biofilm growth occurs and initiates the process of co-aggregation. The co-aggregation is mediated in two ways by the second colonizer. The second colonizer may bind to the specific surface molecules of the biofilms, or the group of bacteria co-ordinates among themselves and favors some phenotypic changes that lead to the co-aggregation on biofilms [7]. The co-aggregation process helps in the extended production of the biofilm matrix and colonizes the maximum surface area of the substrate, protecting from antibiotic stress [30]. One of the common polymicrobial biofilms is observed in acute otitis media, which is associated with 700 million cases reported every year globally [31]. Polymicrobial bacterial biofilms of otitis media, M. catarrhalis, non-typeable H. influenzae, and S. pneumoniae are responsible for the failed or ineffective antibiotic therapy [32]. A common polymicrobial interaction observed between P. aeruginosa and S. aureus in cystic fibrosis lungs and wound infections lead to enhanced severity in patients. It was observed that S. aureus infections are often experienced during childhood, whereas P. aeruginosa colonization increases along with age. P. aeruginosa has an antagonistic relationship with S. aureus. The coexistence and virulence of these versatile pathogens increase upon stress conditions such as antibiotic therapy [33]. 4.2. Bacterial-Fungal Biofilms Candida species are one of the leading opportunistic fungal pathogens responsible for worldwide nosocomial infections. In general, C. albicans biofilms cause infections in immunodeficient patients. Recurrent candidaemia is one of the complications of candida infections, leading to more than 50% of the global mortality rate globally [34,35]. Complex multispecies interactions involving neutral, synergistic, and antagonistic associations are observed with C. albicans on both biotic and abiotic substrates [36]. The fungal-bacterial interactions occur through co-aggregation and adhesion. The adhesions, such as hyphal-associated adhesions of fungi and other adhesins present in the cell surface of bacteria, help in the interactions [37]. Fugal-bacterial biofilms are associated with bloodstream infections, and the treatment strategies focus on anti-fungal and anti-bacterial therapy. C. albicans forms polymicrobial biofilms along with S. aureus and S. epidermidis in nosocomial bloodstream infections and enhances the antibiotic resistance pattern [38]. The polymicrobial interactions of Candida sp. and Streptococci involved in oral candidiasis are synergistic. This association takes place with the help of adhesins present in the hyphal cell wall (Als 3) and bacterial cell surface (SspB). In this context, a bacterial partner assists the fungi to improve the biofilm formation and thereby pathogenesis of Candida by providing nutrients from salivary pellicle. Fungi enhance the Streptococcal growth by supplying nutrients and reducing the oxygen levels in the surrounding areas [39]. 4.3. Fungal-Fungal BiofilmsFungal-fungal mixed biofilm interactions are much less studied. The studies so far conducted have shown active polymicrobial synergistic interactions of fungal-fungal biofilms in infections. The mixed co-infection of C. albicans and C. glabrata showed synergy in developing denture stomatitis. C. albicans help C. glabrata to invade the host epithelium and to establish infection [40]. Other mixed fungal biofilms are between C. albicans and C. dubliniensis, and C. albicans and C. rugosa, which exhibited decreased susceptibility towards anti-fungal agents and enhanced infection rates in the host [41,42]. 6. Model Systems to Study Polymicrobial BiofilmsThere are several model systems developed to study the interactions of polymicrobial biofilms and associated infections. In vitro and in vivo model systems are employed to study polymicrobial infections caused by mixed biofilms and to evaluate antimicrobial susceptibility testing. There is growing evidence showing the developments in model systems used to study polymicrobial infections [98]. The in vitro models include cell-line infection systems, whereas in vivo animal models include rats, mice, etc. Appropriate animal models are essential to understand the complex nature of polymicrobial infections and their treatment methods. The merits of using animals over in vitro models include the presence of an excellent immune system, well-arranged organ systems, availability of numerous molecular reagents to study immune responses, and the genetic background of inbred mouse to decipher the pathogenesis of polymicrobial communities [99]. Table 1 represents different model systems used to study polymicrobial biofilms.These in vitro model systems are cheap, reproducible, and have fewer ethical concerns in studying composition, biofilm formation, and in evaluating anti-microbial testing for polymicrobial communities. There are in vitro static (microtiter plate) and continuous chemostat models to study oral biofilms. The continuous models include chemostat (bioreactor with continuous flow of media and nutrients), flow cell (slides comprising media) and constant-depth film fermenter (fermenter containing coupons suspended from the lid). The modern technology including microfluidics, continuous and static models allow the growth of biofilms [100]. For instance, a costar 24-well flat bottom cell culture plate was employed by Manavathu and co-workers to develop a polymicrobial biofilm model of A. fumigatus and P. aeruginosa. These pathogenic biofilms are observed in the CF lung airways, where they complicate antibiotic treatment. Thus, the in vitro biofilm static model enabled studies of the effect of antibiotics and combinations thereof (cefepime, tobramycin and posaconazole) [101].In vitro models allowed studies of the growth of aerobic and anaerobic bacteria in chronic wounds. Chronic wound biofilms comprise mixtures of aerobic and facultative aerobic pathogens such as P. aeruginosa, E. faecalis, and S. aureus. Lubbock chronic wound biofilm (LCWB) model was used to evaluate the growth of facultative anaerobes in a completely oxygen-rich environment of wounds. The LCWB model provides similar circumstances for in vivo wound biofilms. The in vitro model is enriched with wound-simulating media, oxygen content, nutrients and an insoluble fibrin network provided by S. aureus. This wound model enables the attachment of bacteria and to form biofilms [110]. LCWB is one of the widely used models for studying the effectiveness of many anti-microbial agents. For the in vitro evaluation of anti-microbial dressings, a 48-h viable LCWB wound biofilm model was employed. The LCWB model was transferred into an artificial wound bed containing gelatin and agarose which remained viable for next 48 h and served as an effective pre-clinical in vitro experiment to study polymicrobial biofilms of S. aureus, E. faecalis, B. subtilis, and P. aeruginosa [111].Caenorhabditis elegans is a transparent nematode, another widely used model system due to the ease in maintenance to study molecular and genomic approaches in microbial infections [112]. It serves as an excellent in vivo model to decipher host-pathogen interactions. This model allows the analysis of various virulence factors of pathogens and to study host defence molecules produced upon infection. The pathogenic biofilms initially colonize in the gut, leading to the disruption of the gut lining and eventually organ dysfunction and death [113]. A research group evaluated the role of EPS in the formation of polymicrobial biofilms of S. epidermidis and C. albicans using a C. elegans infection model. The study showed that hyphal formation by fungi and EPS production of bacteria led to increased virulence in the infected nematode and eventually caused death [114].In vivo vertebrate models are most reliable and promising approaches to study polymicrobial biofilms and their interactions with the host. Dalton and colleagues developed a murine wound infection model to study polymicrobial interactions of wound pathogens S. aureus, P. aeruginosa, E. faecalis, and F. magna [57]. The mouse wound model displayed the role of polymicrobial biofilms in delaying wound closure and enhancing antimicrobial resistance. It was observed that polymicrobial wound infections showed improved antibiotic tolerance and delayed wound healing compared to monospecies biofilm interactions [57]. Polymicrobial biofilm formation by P. aeruginosa and B. cenocepacia in CF lungs were demonstrated in CFTIR-deficient mice. The co-infection of bacterial pathogens in mice led to the establishment of polymicrobial biofilms followed by chronic lung infection. The authors studied the effect of interspecies interactions on the development of chronic infections and enhanced production of inflammatory responses in the mice model [115].The polymicrobial consortium of periodontitis consisting of P. gingivalis, T. denticola, and T. forsythia and its synergistic interactions leading to the increased inflammation was studied using a rat model. In the polymicrobial disease model, rats infected with pathogens exhibited chronic inflammation of periodontal structures, causing alveolar bone resorption. Thus, the rat model enabled an evaluation of the combined virulence mechanism of periodontal pathogens leading to chronic inflammation [116]. Likewise, a pneumonia mouse model with polymicrobial infections caused by P. gingivalis and T. denticola led to enhanced respiratory infections. The polymicrobial infections showed a higher mortality rate in the mouse model compared to the monospecies infections. The pathogenic biofilms developed bronchopneumonia and lung abscesses in mice which caused a higher mortality rate and markedly higher production of inflammatory cytokines [117]. The otitis media rat model was used to study the interactions of MRSA and P. aeruginosa in a host. The polymicrobial biofilms were established in the middle ear, causing chronic suppurative otitis media. The colonization of bacteria in the middle ear of rats caused increased production of inflammatory responses that were observed during gene expression studies [118]. Thus, the model systems are highly essential for evaluating the polymicrobial interactions and establishing biofilm infections and host-pathogen responses. 7. Metagenomic Approaches in Detection, Prevention, and Inhibition of Polymicrobial BiofilmsThe current standard approach for polymicrobial detection remains culture-dependent [119]. However, culture-dependent approaches are prone to disadvantages, such as low turnaround and false negatives for slow-growing or low-titer pathogens. Such setbacks can be circumvented by implementing novel molecular techniques such as metagenomics. The advantage of metagenomics is mainly due to its culture-independent and high-throughput nature. Due to its high-throughput nature, metagenomics is widely used in exploring the entire microbial community of environmental or host microbiomes. Metagenomic sequencing can be loosely classified into shotgun or amplicon metagenomics. Although shotgun NGS is the gold standard for metagenomic analysis, 16s rRNA amplicon sequencing is also commonly used. The sensitivity of metagenomic techniques in detecting pathogens in clinical samples has also been demonstrated in several studies. Its advantages over the standard clinical approach include its sensitivity, low sample requirements, identification of pathogens not commonly observed in the standard clinical approach, fast turnaround, and potential to detect novel approaches for infectious disease treatments. The shotgun metagenome approach has several advantages over 16s rRNA metagenomics. It can provide information on the antibiotic resistance mechanism, mutations, metabolic potential, species/strain level information on the taxonomic diversity, and whole genome reconstructions [120]. On the other hand, 16s rRNA metagenomic is cheaper in terms of cost per sample (Figure 2).The human microbiome comprises an intricate microbial network, such as the oral microbiome comprising a complex acidogenic and aciduric microbial community of >700 microbial species, several of which are yet unculturable. Among them, the Lactobacilli and mutans Streptococci (i.e., S. mutans and S. sobrinus) are known to cause caries [121]. The complex interaction between the microbial community has led to the polymicrobial synergy and dysbiosis (PSD) hypothesis, which states that oral disease could be the consequence of dysbiotic microbials rather than a specific pathogen. Similarly, the keystone pathogen hypothesis states that certain low-abundancy pathogens can modulate a healthy microbiome into a dysbiotic microbiome. Polymicrobial natural infections can be viewed based on the interactions between the species. Microbial interactions are often evolutionarily related, and the species within the polymicrobial community interact to protect each other from antibiotics [122]. Metagenomic analysis of such complex microbial communities has provided novel insights into the polymicrobial infections and involvement of keystone pathogens [123]. For instance, millions of diabetic patients are affected with a diabetic foot, characterized by bacterial infections and biofilm. Metagenomic analysis of the diabetic foot identified enrichment of multiple pathogens, including Streptococcus and Corynebacterium, while traditional culturable techniques identified Pseudomonas, Proteus, Enterococcus, and Staphylococcus [124]. Similarly, metagenomic studies of tuberculomas (n = 14) have revealed the lack of M. tuberculosis as dominant taxa in most samples, suggesting the polymicrobial nature [125]. Thus, metagenomic studies have provided novel insights into polymicrobial infection and their interactions mediated by keystone pathogens.The sensitivity of metagenomics has also been demonstrated in the detection of polymicrobial infections in arthroplasty, surgical restoration of joint functions, which is mainly complicated by prosthetic joint infection (PJI). A routine clinical approach for the detection of PJI yields 20–35% false negatives [126]. 16s rRNA amplicon metagenomic screening of PJI though synovial fluid (SF) (n = 22) of PJI patients (n = 11) identified every pathogen that was detected with traditional cultures and also categorized them based on the arthroplasty stages and reduced turnaround time (two days instead of seven days in standard clinical approach). Similarly, the application of 16s rRNA metagenomics in polymicrobial detection has been studied in the sputum of cystic fibrosis (CF) patients with promising results: metagenomics was able to detect a significantly higher number of unique microbes (n = 122), including low-abundance and fastidious microbes compared to the culturable method (n = 18) [127]. Furthermore, the sputum samples could be classified into multiple groups (n = 5), suggesting the implication of a more specific diagnosis. The higher detection rate of pathogens through metagenomics has also been demonstrated through 16s rRNA amplicon metagenomic analysis of brain abscesses (n = 51) which detected more bacterial taxa when compared to the cultural approach with at least two bacterial taxa detected in most of the samples (n = 31), suggesting the polymicrobial nature of the disease [128]. Metagenomic analysis of samples with monomicrobial infections (n = 8) has a detection rate of 100%, but only about 58.2% and 74.5% at species and genus level, respectively, for polymicrobial infections (n = 55) [129]. Metagenomic analysis was also more rapid, with less than 24 h turnaround, and cost-effective compared to the standard clinical approach. The metagenomic approach has also been reported to detect potential pathogenic bacteria that were not detected in the culturable approach and predicted antibiotic susceptibility for 76.5% in the polymicrobial samples [129]. The strength of metagenomics has also been demonstrated in the detection of community-acquired pneumonia (CAP) (n = 59), which was able to detect pathogens not detectable with traditional techniques; some of the patients were detected with polymicrobial infections (n = 31), which led to increased outcomes and reduced the mortality rate [130].Current standard practice for sepsis diagnosis is based on the culture of microbes from the bloodstream. However, the procedure is prone to underestimation or false negatives when the causative agents are in low abundance or difficult to grow. Sepsis can also be the manifestation of the polymicrobial interaction. A recent metagenomic study fortified with machine learning on 287 cohorts demonstrated that polymicrobial sepsis infections in blood-borne infections could be efficiently detected [131]. The authors report interesting findings. The model without the culture-confirmed pathogens performed well, and the model with only single-feature did not perform well, suggesting the polymicrobial nature of the species. Shotgun metagenomic analysis has also been highly efficient in detecting pulmonary infection. For instance, a large-scale metagenomic analysis of patients (n = 235) with suspected pulmonary infections or in a respiratory intensive care unit (RICU) or ventilator identified lower alpha diversity in confirmed patients [132]. A shotgun metagenomic study of periodontitis patients (n = 43) revealed that healthy subjects had higher alpha diversity, and the development of a machine learning model with a naive classifier was able to classify patients and healthy subjects with an accuracy of 94.4% [123]. The authors also determined multiple antibiotic resistance genes and polymicrobial keystone species: P. gingivalis, Haemophilus haemolyticus, Prevotella melaninogenica, and Capnocytophaga ochracea. Such studies provide clear evidence that the implementation of machine learning in metagenomic analysis has promising potential in disease detection and specialized therapeutics based on the nature of the infection.One of the challenges in the shotgun metagenomic is the low sequencing depth of bacterial DNA due to the high load of host DNA. In the shotgun metagenome of human biopsy samples, most reads (>95%) can be attributed to the human host. Hence, a large number of reads are not suitable for the diagnosis. A novel DNA extraction technique was introduced wherein the intact DNA from extracellular or intact human cells is removed: the human cells are lysed in hypotonic treatment followed by digestion of the extracellular DNA with endonuclease digestion [133]. With such selective removal of the host DNA, the resultant reads are substantially high bacterial DNA reads and consequently detect more taxa. Furthermore, despite the sensitivity of the metagenomic approach, some reports have indicated the weakness of metagenomics in comprehensively detecting pathogens [134]. Hence, it is also important to note that some of the microbes detected through the cultivable approach were not detected in the NGS approach. However, the authors note that the de-nosing step in the metagenomic pipeline should be performed with care since closely related sequences are grouped as one, which can lead to loss of genetic diversity in the sample [127]. Hence, metagenomics can be applied in clinical settings as a supplementary approach to the existing cultivable approach. In addition, future directions for polymicrobial detection through metagenomics would include (i) implication of longer reads to obtain longer reliable assembled reads with higher resolution of identification, (ii) reduction of the cost per GB affordable to the clinics in low and middle-income countries, (iii) automatic pipelines friendly to the medical practitioners, (iv) open-source machine learning models in determining infection sub-groups, (v) application of machine learning approaches in denoising, quality control steps, and suggestion of the suitable measures based on the microbiome profiles. Discovery of Novel Biofilm and Quorum Sensing Inhibitors through MetagenomicsBiofilm formation by pathogens is one of the leading causes of antibiotic resistance and protects the pathogens from the host immune system. Thus, biofilm has rendered most antibiotics ineffective. Inhibition of biofilm is one of the main approaches to address microbial infection since ~75% of the microbial pathogens produce biofilms. Quorum sensing (QS) modulates microbial virulence, biofilm, and overall community behaviour. Hence, quorum sensing inhibitors (QSI) are potential agents for modulating biofilm formation and the virulence of the pathogens. N-acyl-homoserine lactones (N-AHLs) are a common class of QS signaling molecules in bacterial cell-cell communication. N-AHLs can be inactivated, also known as quorum quenching (QQ), by enzymes such as N-AHL hydrolase and N-AHL acylases. Recently, quorum quenching activity of oxidoreductases was identified from a soil metagenome [135]. It significantly reduced biofilm formation, pyocyanin production, motility, and the transcription of lasI and rhlI in P. aeruginosa, suggesting its importance in QS and biofilm inhibition strategies.Since metagenomics does not rely on selective enrichment of genes or enrichment of the microbial community, it has the potential to discover novel QQ enzymes. The strength of metagenomics in discovering novel enzymes with QQ activity has been demonstrated in several reports. Novel QSIs such as bpiB01, bpiB04, bpiB07, and aii810 were discovered through a metagenomic approach that encodes for N-acyl-homoserine lactonase and exhibits antibiofilm activity in P. aeruginosa [136,137]. It was stable at below 40 °C, with neutral pH, and attenuated biofilm formation and the virulence of P. aeruginosa. Similarly, a novel hydrolase, 70 kDa BpiB05, that acts on N-AHLs, was identified through metagenomics that is dependent on Ca2+ reducing P. aeruginosa motility, pyocyanin, and biofilm [138]. It has been proposed that marine microbes are a rich source of bioactive molecules with QSI activity [139]. Screening of a metagenomic library constructed from a marine microbial community exhibited a strong QSI activity by ~7% of the clones (out of 2500 clones). The clones were able to disrupt and reduce the QS and biofilm formation in P. aeruginosa and A. baumannii [140].Small peptides are a potential agents for biofilm inhibition. However, experimental screening of thousands of peptides is costly and resource-intensive due to the limited number of peptides with antibiofilm properties. In this context, prediction models based on the Support Vector Machine (SVM) approach have been attempted, with promising results [141]. Similarly, efforts to extract bioactive peptides from metagenomes also show great potential. Features from the primary structure, physicochemical features, and NMR spectra are fed into various algorithms such as random forest and SVM [142]. 8. Innovative Approaches to Mitigate Polymicrobial BiofilmsThere are several methods or agents that inhibit the formation of biofilms by targeting different biofilm developmental stages. Several novel methods or anti-biofilm agents that are under development can serve as alternatives to antibiotic treatment to reduce the emergence of antibiotic-resistant strains in future [143] (Figure 3; Table 2). As biofilms are highly resistant to antibiotics and fail to reach the deeper layers of biofilms, alternative methods which can degrade biofilm matrix and destroy the deeper resistant cells are recommended [144]. Antimicrobial resistance is often observed in polymicrobial biofilms owing to the presence of antibiotic-resistance genes, interspecies genetic exchange/transfer, production of microbial metabolites and quorum sensing signaling, leading to the altered antibiotic susceptibility. Therefore, multiple mechanisms are adopted by polymicrobial biofilms to promote antibiotic resistance [145]. For instance, transfer or exchange of antibiotic-resistant genes among polymicrobial communities often favours the enhanced antibiotic resistance. There is evidence of horizontal gene transfer (HGT) among mixed cultures of methicillin-resistant S. aureus (MRSA) and E. faecium. In mixed cultures, the presence of vancomycin (vanA) and tetracycline (tetU) resistance genes were detected in MRSA, yet had not been present previously [146]. Conjugation and/or transformation are also involved in the transfer of resistance genes in polymicrobial cultures. In a study, the transfer of tetracycline (tetM)-resistant genes to Streptococcus sp. was observed either by conjugation or transformation with Veillonella dispar [147].Studies showed polymicrobial biofilms of S. aureus and C. albicans are more recalcitrant to antibiotic treatment regimes. In particular, the polymicrobial biofilms showed 10 to 100-fold resistance to rifampicin, ciprofloxacin, vancomycin, oxacillin and delafloxacin in comparison to the monotherapy in sessile S. aureus cells. The increased antibiotic resistance profile is correlated to the formation of nutrient-deficient persister cells in the biofilms [148].There are enzymes or factors encoded by the bacteria to inactivate or degrade antibiotics. One such example is beta-lactamases, produced by a group of bacteria to inactivate cell wall synthesis inhibiting β-lactam antibiotics. It has been observed that these enzymes not only help their own bacterium which produces the enzyme but also protects the neighbouring bacteria in the polymicrobial interactions [149,150]. Beta-lactamases produced by otopathogen M. catarrhalis confer resistance to beta lactam drugs; other pathogens that produce beta-lactamases are H. influenzae and S. pneumoniae [145,151]. There are several reports of polymicrobial biofilms of bacteria and fungi which showed decreased vulnerability towards different generations of antibiotics. The growing challenge of antibiotic resistance in polymicrobial biofilms causing chronic and clinical infections recommends alternative and effective therapeutic modalities. New anti-biofilm molecules, compounds targeting interspecies interactions/quorum sensing, and anti-biofilm agents combined with commonly available antibiotics are under investigation and will be available in the near future [152]. 8.1. Nanoparticle and Nanoconjugate Mediated TherapyNanotechnology deals with particles of 10 to 100 nm size widely used in different fields such as medicine and dentistry. The unique features of nanoparticles enable them to be used as effective and alternative agents in the treatment of infectious diseases and as drug delivery agents. They have a small size, high surface to volume ratio, chemical and biological reactivity [167]. There are metallic nanoparticles, polymer nanoparticles and nanoparticles with antimicrobial coatings used effectively in the management of infections. Metallic nanoparticles have the ability to interact with microbial membranes, nucleic acids, and proteins, resulting in the anti-microbial activity [168].Nanoparticles exhibit enhanced bioavailability and targeted delivery of drugs to biofilms. There are effective drug delivery nanoparticles made of lipids, silica and polymers. These delivery agents can safely deliver drugs to the target site without being affected by bacterial-deactivating enzymes such as β-lactamases [169]. Metallic nanoparticles such as silver, gold, iron, copper, etc., can act as anti-biofilm agents [170]. Silver nanoparticles (AgNPs) are widely explored anti-bacterial and anti-fungal agents. Yasinta et al., 2021 studied the ability of AgNPs to inhibit polymicrobial biofilms of E. coli and C. albicans. The synthesized silver nanoparticles entered the EPS matrix and degraded the extracellular matrix growth, leading to the destruction of polymicrobial biofilms. Thus, AgNPs showed dual action by degrading EPS matrix and killing the polymicrobial biofilms of E. coli and C. albicans [171].Positively charged silver nanoparticles inhibited single-species biofilms and polymicrobial biofilms of MRSA and C. albicans. AgNPs inhibited the MRSA/C. albicans biofilms in a dose-dependent manner, substantiating the role in inhibition of most of hospital-acquired infections by these biofilms. MRSA/C. albicans biofilms are found in indwelling devices, causing blood-stream infections. In addition, authors explored AgNPs coated catheters to prevent the formation of polymicrobial biofilms and provided evidence of charged silver nanoparticles and nanoparticle-coated catheters in the inhibition of polymicrobial biofilms of MRSA/C. albicans [154]. Along with silver, gold nanoparticles (AuNPs) were also explored as anti-biofilm agents. Interestingly, hybrid nanoparticles were prepared using silver and gold to evaluate their efficacy towards polymicrobial biofilms. Hybrid nanoparticles at very low concentrations inhibited the biofilm formation in P. aeruginosa and S. aureus. Moreover, the mechanism of action towards polymicrobial biofilm was found to be the production of intracellular reactive oxygen species (ROS) [172].Curcumin is one of the phytochemicals which showed an anti-biofilm effect against a broad spectrum of pathogens. Researchers studied the role of curcumin-loaded chitosan nanoparticles against monospecies and polymicrobial biofilms of C. albicans and S. aureus. Chitosan nanoparticles work as a good drug delivery agent for hydrophobic curcumin to the biofilm matrix and favours sustained delivery of drug. Thus, nanoparticles improve the therapeutic effect of curcumin when loaded onto chitosan nanoparticles, which was confirmed by a reduced ability to form polymicrobial biofilms. Curcumin-loaded nanoparticles reduced the biofilm matrix thickness and favoured microbial death [153]. Maxillofacial silicone prostheses are employed in the therapy of head and neck defects. Monospecies or mixed biofilms are one of the reasons contributing to the degradation of these prostheses. Thus, silicone prostheses coated with silver nanoparticles inhibited the bacterial-fungal biofilm formation in vitro. The release of silver ions and cellular permeabilization led to the inhibition of polymicrobial biofilm formation by S. aureus and C. albicans [173]. Thus, the studies explored the opportunities of using nanosized particles to treat polymicrobial biofilms and to control the emergence of antibiotic-resistant strains. 8.2. Antimicrobial Photodynamic Therapy (aPDT)Antimicrobial photodynamic therapy is an emerging and effective therapeutic strategy to treat acute and chronic infections caused by planktonic cells and biofilm pathogens. aPDT is a light-dependent treatment method which involves the production of reactive oxygen species through type 1 or type 2 pathways [174]. In aPDT, a non-toxic and light-sensitive compound termed a photosensitiser (PS) in its excited state interacts with molecular oxygen surrounding the cells and produces ROS such as hydroxyl free radicals, peroxides, superoxides and singlet oxygen [175]. ROS can act on different cellular targets such as proteins, DNA and lipids, thus potentiating the non-specific action of aPDT towards the elimination of resistant strains [176]. The role of PDT in the treatment of broad-spectrum biofilm pathogens was studied in both in vitro and in vivo conditions. Several studies are in progress with pre-clinical and clinical trials to evaluate the effectiveness of aPDT [177,178,179]. The mechanism of action of aPDT against polymicrobial biofilms is provided in the Figure 4.Polymicrobial Gram-positive and Gram-negative pathogens are associated with the failure of endodontic treatments. Methylene blue (MB) is one of the widely used anti-microbials (PS) and showed effective phototoxicity against polymicrobial pathogens causing endodontic infections. Methylene blue (10 µg/mL) mediated aPDT reduced polymicrobial biofilms comprising Actinomyces israelii, F. nucleatum subspecies nucleatum, P. gingivalis, and P. intermedia on root canals. Thus, aPDT is an excellent alternative for the treatment of root canal infections [180]. Toluidine blue O (TBO) is an excellent anti-bacterial cationic PS used for broad-spectrum activity [181]. Akhtar et al. 2021 employed a novel nano-phototheranostic approach to prevent polymicrobial and monomicrobial biofilms of diabetic foot ulcer. In this method, anti-microbial agents, silver and gold nanoparticle cores, were coated with chitosan and conjugated with TBO to enhance the overall aPDT mechanism. The synthesized and photoactivated nano-PS conjugate reduced both monomicrobial and polymicrobial biofilms containing two versatile diabetic foot ulcer-causing pathogens, P. aeruginosa and S. aureus [182]. Similarly, nanoparticles were conjugated with TBO for enhanced loading and maximum phototoxicity against wound polymicrobial biofilms. Anionic surfactant dioctyl sodium sulfosuccinate-alginate nanoparticles were prepared and loaded with TBO and used against planktonic and mixed biofilms of MRSA and P.
留言 (0)