
記住我
The models referred to in this review encompass a range of mouse and rat strains. Unfortunately, some of the studies included use models on a background of the C57BL/6 mouse strain, which is known to be associated with a peripheral, progressive high-frequency sensorineural hearing loss from 3 months of age, and is even used as a model of early onset hearing loss [95]. Auditory experiments conducted in these animals therefore have an underlying caveat affecting wild type (WT) hearing, which may make it difficult to parse out differences in audition caused by the autism model versus those resulting from the strain’s genetic background. There were 17 studies that used this mouse strain (above 3 months of age) and this is noted throughout this ‘Results’ section, as well as in Supplementary Table 1.
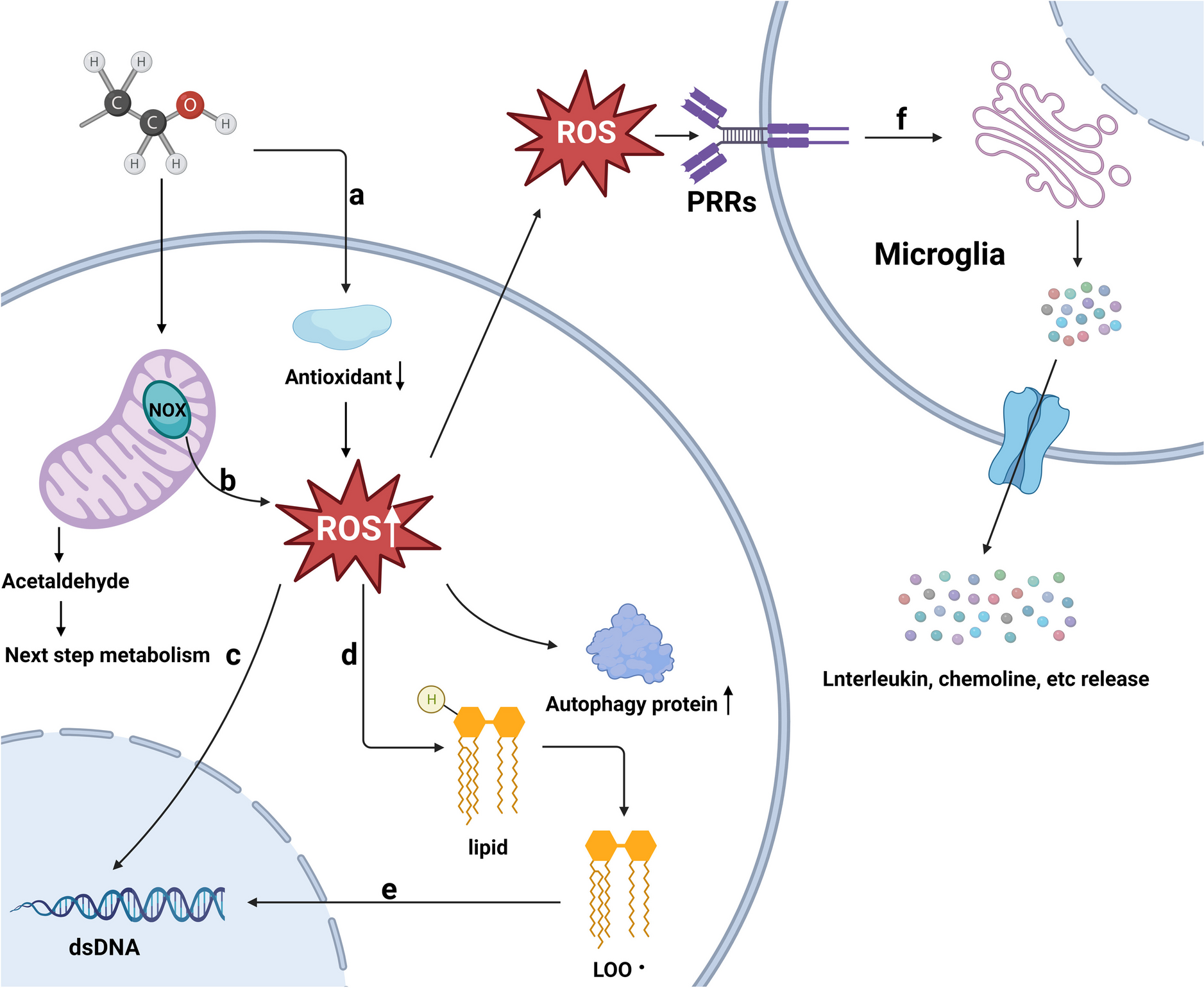
Non-invasive measures of auditory functionAuditory brainstem responsesAuditory brainstem response (ABR) recordings are non-invasive measures used clinically by audiologists to assess the transmission of information through the subcortical auditory pathways and intensity thresholds for responses to sound [51]. Equivalent recordings to those in humans of all ages can also be made in rodents, where they are often used to measure auditory function, particularly auditory thresholds (sensitivity). The output trace has several peaks, which correspond to the activity in subsequent brain regions along the auditory pathway in response to a sound stimulus (Fig. 3A). In children with autism, the most common findings from ABR recordings are increased latencies, especially of peaks III and V (representing delayed activation of the SOC and IC), although this effect is reduced or even reversed in adulthood [82, 99, 126]. Findings of changes to the amplitude of ABR peaks (which would correspond to more or less activity in the relevant brain area) in autism are much less conclusive [99, 126]. Results of ABR studies in rodent models of ASD are summarised in Table 1. The majority of studies show no difference in ABR thresholds compared to WTs, although half of these studies were conducted in C57BL/6 mice over 3 months old [37, 64, 71, 89, 124, 152]. However, Fmr1−/− and Adnp+/− mice have higher thresholds than WTs, indicating less sensitivity to quieter sounds [50, 116]. A study in 16p11.2 microdeletion mice also used ABRs to test hearing thresholds, but found no responses and concluded that this mutation caused deafness in these mice [151].
Fig. 3
Measures of function in along the auditory pathway. A Example human auditory brainstem response (ABR) trace. The first peak represents activity in the auditory nerve, and the second correlates to the cochlear nucleus. Peak III represents the superior olivary complex, and peaks IV and V represent the lateral lemniscus and the inferior colliculus, respectively. B Example cortical auditory event-related potential (ERP) trace. The ERP is made up of 5 peaks, representing activation in different areas. P1 is produced by activity in the auditory thalamus (MGN) and the primary auditory cortex, N1 is produced by the auditory cortex. P2 is produced by the association cortex. N2 and P3 peaks (associated with the frontal cortex) are observed in humans, but less commonly in rodents. C Exemplar EEG frequency activity. Raw EEG traces (as may be recorded from the auditory or prefrontal cortices) are composite waves, from which activity in several frequency ranges can be extracted. The power in each of these frequency bands indicates the extent to which activity at that frequency contributes to the overall recorded activity
Table 1 Auditory brainstem responses in rodent models of ASD. Model names are written in bold if the result represents more than one study, or the majority of studies in the case where results from a single model are mixed. Numbers in subscript brackets refer to the relevant papers from the literature search, as numbered in Supplementary Table 1. Models are listed in alphabetical orderChanges in the amplitude or latency of the ABR peaks in autism models would indicate changes in the intensity or delay of the response in certain auditory structures. However, studies of the amplitude and latency of ABR peaks in autism models are sparse and have inconclusive results (Table 1). Most studies show no difference to WTs, and in several cases where there are differences at young ages, these shift to resemble WTs in older animals [71, 116, 124]. Differences in ABR latencies are also most evident in humans at young ages, and it has been recommended that rodent studies examine a range of ages in order to capture such developmental changes between juveniles and adults [128]. Most studies using ABRs have found few and varied differences between WTs and ASD model animals, and there are few coherent patterns across studies within and across rodent models. As appealing as ABRs are as a non-invasive measure of function in lower auditory structures, too few studies have performed this test on rodent models of ASD to draw conclusions across different models. ABR measures in children with autism show trends towards lower amplitudes and longer latencies, but there is a significant variation in the response [74, 76, 136]. The inability of rodent studies in various models to converge on a consistent ABR phenotype may therefore be an accurate portrayal of the diversity of ABRs in iASD with a range of aetiologies.
Cortical event-related potentialsEEG is a useful non-invasive tool for measuring cortical activity in humans, both at rest and in response to stimuli. In rodents, EEG generally uses implanted electrodes, and typically just one to three sites, though more recent studies have used up to 30 channels [55]. These methods can be used to measure the cortical auditory event potentials that follow ABRs in response to sound stimuli (Fig. 3B). As the included studies refer to these waveforms simply as auditory event-related potentials (ERPs), that term will be used in this review. These ERPs have a distinct shape, including well-defined positive peaks (P1, P2, and P3) and negative peaks (N1 and N2), which are associated with different aspects of sound processing [131, 148]. The P1 is produced by a combination of activity in the auditory thalamus and the primary auditory cortex, the N1 by the auditory cortex, and the P2 by the association cortex [59, 83, 148]. The N2 and P3 peaks (associated with the frontal cortex) represent higher-order activity and are observed in humans, but appeared in very few of the studies included in this review. The N1 peak has been shown to be reduced in amplitude in people with autism, but increased in amplitude in those with FXS [35, 83, 107, 131]. These changes represent reduced evoked activity in the auditory cortex in iASD, but stronger activity in the auditory cortex in FXS. The N1 amplitude is therefore a prime example in which auditory phenotype is certainly not generalisable across aetiologies. In Rett syndrome, the amplitude of N1, P2, and N2 may be decreased, and latencies across most peaks tend to be increased [119, 136]. In people with iASD, the most consistent difference in the ERP profile is increased latency of the N1 peak, indicating slower transmission of information to the auditory cortex [43, 105]. It is known that ERPs change with age in humans and rodents, notably that the N1 and P2 amplitudes are lower, and P1 and N1 latencies are increased in children and young mice compared to adults [131]. The reduced N1 amplitude and increased N1 latency seen in people with autism may therefore represent delayed maturation of auditory circuits.
A summary of analyses of auditory event-related potentials in rodent models of ASD is shown in Table 2. In cases where more than one study examined the same model, the most common result is indicated by bold font. It should be noted that a subset of these studies measured ERPs in anaesthetised animals [28, 30, 32, 60, 70, 123]. Anaesthesia has been shown to disrupt auditory and other sensory event-related potentials [5, 114]. Results in Table 2 which come solely from studies in anaesthetised animals are therefore indicated with an asterisk. It is noted that these studies are largely in rats: only two Fmr1−/− mouse studies used anaesthesia, and the results from these merged with those from awake Fmr1−/− mice. The majority of studies used white noise as a stimulus, though some used recorded human speech sounds or pure tones [28,29,30, 32, 60]. Human studies typically use simple stimuli such as pure tones, but it has been suggested that more complex sound stimuli can be informative and should also be used in rodent studies for translational validity [83, 125]. Furthermore, most studies do not compare a range of stimulus presentation rates to generate average ERP traces. One study that did so found the habituation rate of the N1 amplitude was lower in Fmr1−/− mice, but only at presentation rates greater than one stimulus per second [70]. This difference in habituation could lead to detection of differences in N1 amplitude in the mean trace.
Table 2 Auditory event-related potentials in rodent models of ASD. Entries are in bold if they are supported by more than one study or represent the result of the majority of the studies using that model (if there is no majority, no result is in bold). Numbers in subscript brackets refer to the relevant papers from the literature search, as numbered in Supplementary Table 1. Asterisks indicate cases where all of the studies contributing to that result were conducted in anaesthetised animals. Models are listed in alphabetical order P1Many studies with Fmr1−/− mice have found no difference in the amplitude of the P1 peak, though a recent study using a higher number of channels found an increased P1 in Fmr1−/− mice compared to WTs [55, 60, 67, 130, 147]. The P1 amplitude is generally not different to WTs in other ASD models, with the exception of 15q13.3 deletion mice [48, 57, 64, 121].
N1In Fmr1−/− mice, the amplitude of N1 is usually increased compared to WTs, more commonly when measured from the frontal rather than the auditory cortex [55, 60, 67, 70, 130, 147]. However, N1 amplitude is decreased across the auditory cortex in anaesthetised Fmr1−/− rats [30]. The increase seen in the mice resembles the auditory endophenotype of people with FXS. The amplitude of the N1 peak in other models is variously increased, decreased, or unchanged [8, 20, 28, 32, 48, 57, 64, 106, 121, 123]. Some of the increased and unchanged N1 amplitude results [8, 57, 64, 106] come from studies in C57BL/6 mice over 3 months old, which may impede their ability to accurately portray an auditory endophenotype. Nevertheless, it appears that the consistently reduced N1 amplitude in people with iASD and Rett syndrome is not well recapitulated across a wide range of rodent models.
P2Mixed results are also visible in the amplitude of the P2 peak, with awake mice again mostly showing increased or unchanged amplitudes [48,
留言 (0)