
記住我
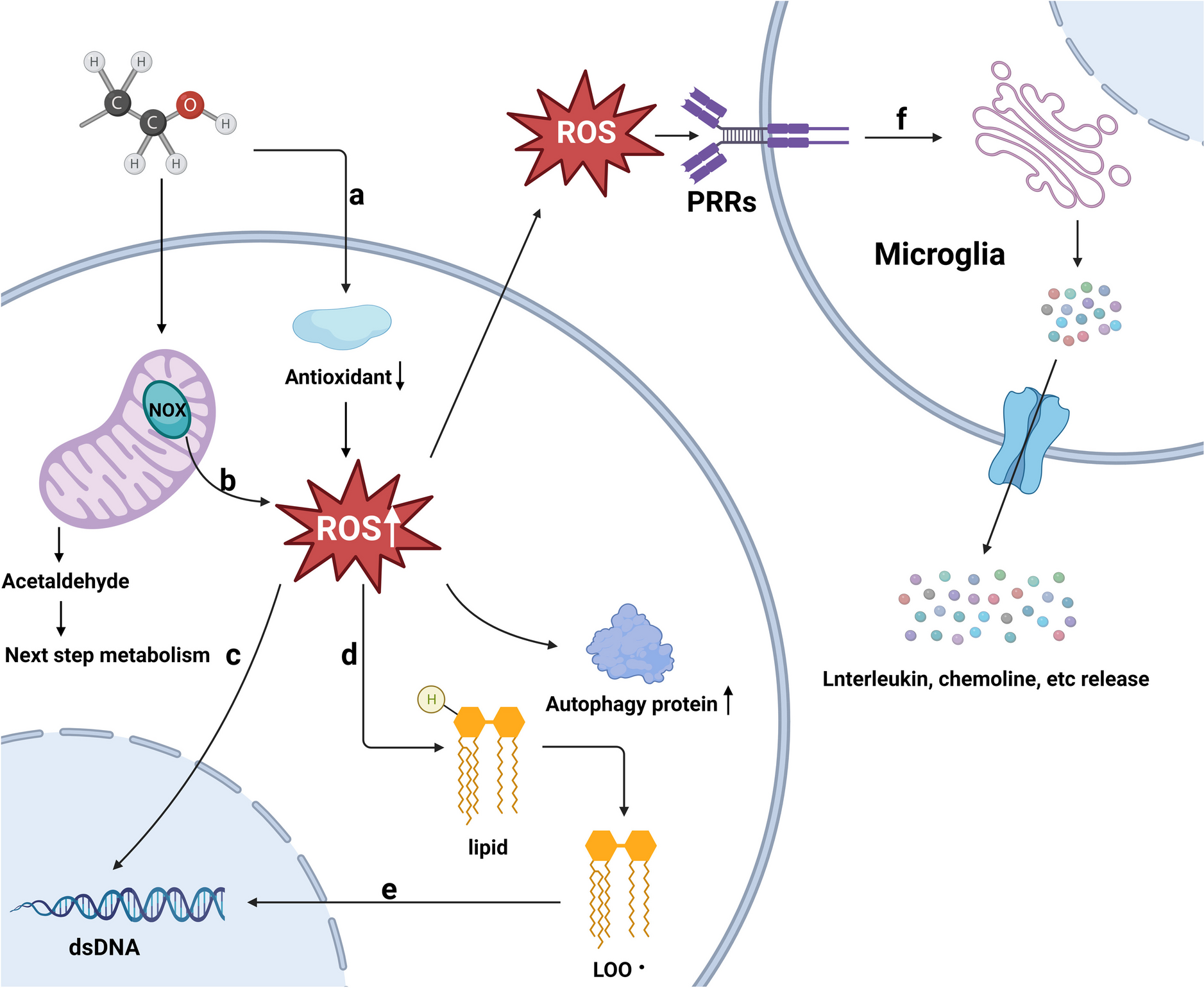
We recruited two groups of participants, NT and cognitively able autistic adults. We choose a male-only sample, since ASD is diagnosed with a 4.3 times higher rate for boys than for girls [39]. All participants reported normal or corrected-to-normal vision and had more than 9 years of formal education. Cognitively able autistic participants scored above 70 points in the verbal subscale of the Wechsler Adult Intelligence Scale and had been diagnosed with ASD (based on DSM-V criteria of the American Psychiatric Association, [2]. Additionally, we used the Asperger’s Syndrome Diagnostic Scale (ASDS;[45]) to confirm the clinical evaluation diagnosis and to characterize the level of autistic traits in this group (see Table 1). The study was conducted according to the Declaration of Helsinki [70] and was approved by the ethical review board of the Faculty of Psychology at the University of Lisbon. All participants gave their written consent after being informed about the procedure and were given the opportunity to ask questions. The data from one cognitively able autistic participant were excluded from further analyses due to low performance (d′ lower than 2 SD below group mean), and the data from one NT adult were excluded due to extensive artifacts (81.7% of all trials had to be removed). The two groups were matched for age, schooling, and general cognitive abilities (assessed with Raven’s progressive matrices) after excluding 4 NT adults with extreme values in age and general cognitive abilities. Thus, the data of 17 NT and 14 cognitively able autistic participants were included in the analysis (for details, see Table 1).
Table 1 IQ and demographic information of both groups of male participants, indicated separately for cognitively able autistic and neurotypical (NT) adultsMaterials and procedureStimuli were 1600 color photographs; 400 animals and 400 food items were used as targets and 800 objects as non-targets. Targets and non-targets were presented in random order with equal probability (50%). In each trial, one picture of one of these categories was presented. None of these images is generally suspected of producing special emotional arousal. Of course, we cannot exclude such an effect on an individual level. Stimuli were not controlled for color parameters or visual complexity. Throughout the experimental blocks, a black fixation frame at the center of a 10° × 10° visual angle (as described in [67] was visible on a CRT monitor with a refresh rate of 85 Hz and a resolution of 1024 × 768 pixel. Each picture was presented only once in a standardized size (10° × 10° visual angle). In a prior rating test, 17 NT students (mean age = 19.4 years; 2 males) were asked to indicate how well each item represented a given category on a 7-point scale, i.e., “animal” or “food,” respectively. Note that we used an independent sample to rate typicality. The z-transformed means served to define typicality level; values below 0 were used as atypical items and those above 0 as typical items. Based on this definition, the mean original typicality ratings (1–7) were significantly different for both categories (Fig. 1). Hence, 200 images were typical, and 200 images were atypical for each category (i.e., 200 typical animal, 200 atypical animal, 200 typical food, and 200 atypical food images). Stimuli were presented with presentation software (version 18.0, Neurobehavioral Systems). Participants were seated in front of a computer screen, placed at eye level and at an average distance of 40 cm on a table in a dimly lit and shielded room. Participants rested their index fingers on the F and J keys of a QWERTY keyboard and responded by pressing one of the keys. Participants performed a visual categorization task, in which they indicated, for each item, whether or not it belonged to the animal or food category, respectively. Each relevant category was presented in blocks, and the order of these blocks and the response keys (yes, no) were counterbalanced.
Fig. 1
Results of prior rating test. Mean typicality rating of students (n = 17) after dividing images in each category (animals in red, food in blue). The asterisk indicates a significant difference between typical and atypical items in both categories
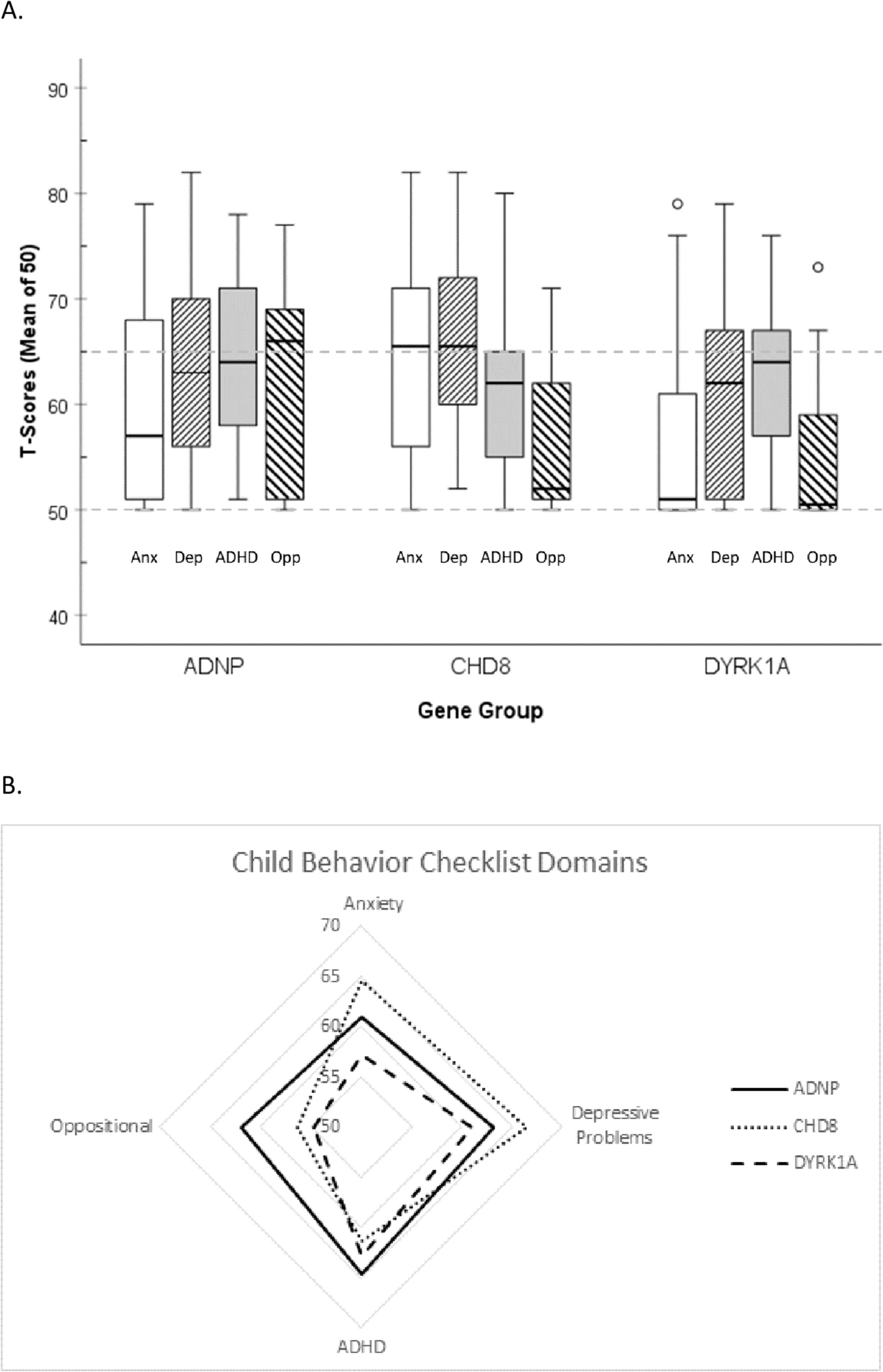
Presentation times (short or long) were varied randomly, with equal proportions of targets and non-targets. Stimuli were presented for 2 frames (23.5 ms) in the short condition and for 7 frames (82.3 ms) in the longer presentation duration. Each subsequent stimulus was presented after a random interval between 200 and 500 ms following the response, with a maximum of 1500 ms (displayed in Fig. 2). After every 100 trials, a short break was offered.
Fig. 2
Trial design. For the whole trial and during each block, a fix square was displayed. In each trial, images were presented in the square for either 23.5 or 82.3 ms. Participants could respond until 1500 ms after stimulus offset to a two-option forced choice (“yes”/”no”), answering the question “Did you see an animal image?” Participants were asked to respond as accurately and as fast as possible
EEG recordingFor the EEG recording, we used 64 Ag/AgCl cap-mounted electrodes, plus two placed at the mastoids and four around the eyes, positioned on an extended 10–20 system [25]. The EEG was recorded with the BioSemi EEG-System (BioSemi B.V., Amsterdam, Netherlands). All electrodes were recorded with an electrode offset within a 40-μV range. The electrode offset is generated at the junction of the skin and electrolyte solution under the electrodes. It is a by-product of the direct current potentials and results in a voltage at the amplifier input (Jones, 2015). We used the electrodes around the eyes (above and below the right eye and beside the right and left eye) to record eye movements. The ground electrode was placed with the common mode sense (CMS) active electrode and the driven right leg (DRL) passive electrode at the electrode positions PO1 and PO2, respectively, in the 10–20 system. The CMS is also used as online reference. The sampling frequency was 2048 Hz. EEG signal was filtered online with a 0.16 Hz high-pass filter and a 100 Hz low-pass filter.
EEG data processingWe used the spherical spline method [50] for interpolation of electrodes with many artifacts, since this method makes no assumption about the conductivity of the head tissues [9],on average in NT adults: 1.9 electrodes; in cognitively able autistic adults: 2.6 electrodes, ranging for both groups between 0 and 8 interpolated electrodes). The signal was re-referenced offline to the average of all cap-mounted electrodes using BrainVision Analyzer 2.1 (Brain Products GmbH, Gilching, Germany). The choice of reference depends on a variety of factors, including the number of electrodes, location of electrodes, cognitive task, analyses to be performed, and brain regions to be investigated (see [9]. Since most of the relevant portions of ERPs in cognitive neuroscience consist of frequencies between 0.01 and 30 Hz [35], the EEG signal was filtered using a zero-phase shift Butterworth filter (most common used filter [9], with a high cutoff at 30 Hz at 48 dB/oct. We corrected for eye movement artifacts by using an independent component analysis (ICA) with the infomax-restricted algorithm [28], where possible. For the ICA, we selected a 100 s interval from the 16th block of the experiment as a training data set for computing the unmixing matrix. ICA components were automatically identified by picking up blinks and saccades, as evidenced by their characteristic shape and maximum at frontal sites. After removing these components, the EEG was reconstructed. Two cognitively able autistic participants did not blink during most of the blocks,thus, a suitable amount of data for the ICA was not available [23]. Therefore, we manually deleted all blinks for these participants, which amounted to 38 and 49 blinks, resulting in 1.38 % and 2.21 % deleted trials, respectively. EEG segments were based on a time window of 200 ms before and 800 ms after stimulus onset. Artifacts were removed automatically when (1) the amplitude difference between two sample points exceeds 50 μV, (2) the amplitude difference was more than 150 μV in an interval of 100 ms, or (3) a low amplitude of 0.5 µV occurred in a 100 ms interval [9]. Due to the artifact rejection, on average, 2.48 % of all trials had to be removed in the NT group and 5.07 % in the cognitively able autistic group. On average, there were 114 (range: 49–197) trials left per condition per participant for the NT participants and 109.9 (range: 33–196) for the cognitively able autistic participants (for more details, see Table 2).
Table 2 Trials per condition. Mean number of trials and range, per condition, used in the ERP analysis, calculated for cognitively able autistic adults and neurotypical (NT) adults separatelyAnalysis of behavioral dataCategory discrimination (d′) and reaction times (RTs) were analyzed. For both, responses faster than 200 ms were excluded, resulting in the exclusion of 2.82 % of all trials in the NT group and 2.64 % in the cognitively able autistic group. Statistical analyses were restricted to responses to the target category. RT analyses were based on correct answers only; hence, an average of 10.29 % of all trials had to be excluded in the NT group and 9.64 % in the cognitively able autistic group due to incorrect responses.
Category discrimination was analyzed using d prime (d′). D′ was calculated by using the z score of the probability of hits minus the z score of the probability of false alarms. In the absence of false alarms or misses, we applied the formula by Macmillan and Creelman [37]. A d′ of 0 indicates no discrimination, whereas a d′ of 4 indicates nearly perfect performance [38]. For both mean d′ and mean RTs, we used a repeated measure analysis of variance (ANOVA) with category (animal vs. food), typicality (typical vs. atypical), and presentation time (23.5 vs. 82.3 ms) as within subject factors and group (NT vs. autistic participants) as between subject factor. For the sake of brevity and to ease readability, we report only those effects and interactions with p-values below the conventional significance value of .05; all remaining analyses are not listed in the result section. All recorded p-values were Greenhouse–Geisser corrected, when needed [22].
Analysis of EEG dataA baseline correction was applied to the segmented signal, using the time window of 200 ms before stimulus onset (as recommend by [35]). The signal was averaged per condition and participant. For the first part of the EEG analysis, we compared the averaged signal of correct responses to target and distractor images to replicate the results by Thorpe et al. [64]. We also followed the statistical approach used in that study, i.e., we tested when the onset of the differential activity (targets–non-targets), dN150, diverges from 0 (15 consecutive t-test values below p < .01; [57] as used by [64]). Since Thorpe et al. [64] recorded with a sampling rate of 1000 Hz, we downsampled our EEG signal to this value. Similar to Thorpe et al. [64, 67], we grouped the signal of seven electrodes (Fp1, Fp2, F3, F4, F7, F8, and Fz) for a frontal region of interest (ROI) and six electrodes (O1, O2, Oz, PO7, PO8, and POz) for an occipital ROI for the short (23.5 ms) and the long (82.3 ms) presentation duration with Python 3.6.5. To calculate the peak latency of the differential activity, we selected individual peaks occurring between 150 and 200 ms, for each both ROIs for each presentation durations, with BrainVision Analyzer. To assess whether the peak latencies differed between conditions in each group, we used a repeated measure ANOVA with two variables of two levels, ROI (frontal vs. occipital), category (animal vs. food), and presentation duration (23.5 vs. 82.3 ms) with SPSS 26. To increase readability, we report only main or interaction effects involving the factors category or presentation duration.
In the second part of the EEG analysis, we focused on the N1, P2, N2, and P3 components based on the literature. For these analyses, we only used target stimuli to which participants responded correctly with a minimum RT of 200 ms, in line with the criteria used for behavioral data. These effects were evaluated at pre-specified ROIs according to the literature. The N1 component peaks at around 140 ms after stimulus onset; therefore, we chose a time window between 120 and 170 ms (similar to [52]) to calculate the peak latency and mean amplitude. The ROI for this analysis comprised the average signal of F3, F4, Fz, C3, C4, and Cz (similar to [3]). We calculated the peak latency and mean amplitude for the anterior P2 between 180 and 240 ms (similar to [8]), at an anterior ROI including F1, F2, Fz, FC1, FC2, FC5, FC6, FCz, C3, C4, C5, C6, and Cz [8]. For the anterior N2, we used the time window between 240 and 300 ms (similar to [8]) with a ROI containing Fpz, Fp1, Fp2, AFz, AF3, AF4, Fz, F1, and F2 [40]. The P3 was analyzed between 300 and 500 ms after stimulus onset [52] at a central-parietal ROI (averaged P1, P2, Pz, CP1, CP2, CPz, C1, C2, and Cz, similar to [52]). To assess the peak latency, we used local maxima/minima to identify the peaks in the pre-specified ROI and time window of each ERP component for each participant. This procedure identifies the latency corresponding to the largest/smallest amplitude value in the specified time window and ROI.
For mean amplitudes, an omnibus ANOVA with the factors Group, ROI, Category, Typicality, and Presentation Time for the entire time window of interest (150–500 ms) revealed a significant 5-way interaction (p < .05). A corresponding omnibus ANOVA with the factors Group, Component, Category, Typicality, and Presentation Time for peak latencies revealed another significant 5-way interaction (p < .05), demonstrating the complex interplay of all the factors under investigation for both types of dependent variables. These interaction effects were followed up by performing hierarchical ERP analyses by group, since this approach is required to account for the specifics of event-related potentials from a theoretical perspective; event-related potentials assess the average EEG activity across many trials in order to enhance systematic activity associated specifically with cognitive processing related to a stimulus. This approach effectively reduces the impact of unsystematic fluctuations in the ongoing EEG (i.e., improving the signal-to-noise ratio). However, this approach does not control for factors unrelated to cognitive activity that do not vary across trials (i.e., morphological differences between individuals like skull thickness or myelination) but still can have considerable influence on amplitude differences as measured on the skull. Therefore, comparing microvolt differences in amplitude between individuals does not allow to make inferences about the underlying cognitive processes (c.f. [41, 65]). Moreover, numerous neuroanatomical differences have been described for autistic as compared to NT individuals (for review, see [1]), for instance, differences in total brain volume, regional gray — white matter differences, differences in stacking of neuronal cell bodies (particularly in layers III and V of the neocortex), and differences in the density of the cerebellum and the size of the amygdala [1]. Additionally, anomalies have been found in the biosynthesis and transmission [4, 15,16,17, 46,47,48, 53, 60, 72] as well as expression [10, 56] of the excitatory neurotransmitter glutamate and the inhibitory neurotransmitter gamma-aminobutyric acid in autistic individuals. As these differences in morphology are unlikely to be randomly distributed, EEG waveforms vary systematically in morphology across participant groups, and even with the poor spatial resolution of the surface EEG, these effects are not minimized. Hence, ERP analyses were performed for each group separately, as statistical interactions between groups of participants may reflect anatomical differences rather than different cognitive processes employed in each group.
For each component, we used a repeated measure ANOVA with the following factors: Category (animal vs. food), Typicality (typical vs. atypical), and Presentation Time (23.5 vs. 82.3 ms) for each group. Similar to behavioral analyses, we report effects with p-values below the conventional significance criterion of .05. This also includes significant interaction effects that did not reveal any significant differences between conditions in Bonferroni-corrected post hoc comparisons. In addition, we report all results of special interest (i.e., those explicitly based on the hypotheses or effects that reach statistical significance in one group, but not in the other.) We illustrate the results of these ERP analyses in the figures, focusing on significant main or interaction effects. After the data were extracted for statistical analyses, a 20 Hz high cutoff filter (at 48 dB/oct) was applied to increase readability of the figures. Figures were created in the BrainVision Analyzer and were edited with CorelDraw X7 (Corel GmbH, München, Germany). For the ERP figures, we edited the following aspects in CorelDraw X7: color and thickness of the ERP waves, thickness of the x- and y-axis, font size of the values on the x- and y-axis, added either a gray square to indicate the time window or arrows to indicate the peak an analysis was performed on, adding asterisk to indicate significant differences, added legends, and crop the width of the whole figure at appropriate millisecond to represent the ERP results. For the topographical maps, we added black dots for the electrodes included in the ROI and increased the numbers on the legend in CorelDraw X7.
留言 (0)